Populations and Evolution
arXiv:q-bio.PE
Population dynamics, spatio-temporal patterns, evolution, co-evolution, biodiversity, phylogeny, models of epidemics.
Population dynamics, spatio-temporal patterns, evolution, co-evolution, biodiversity, phylogeny, models of epidemics.
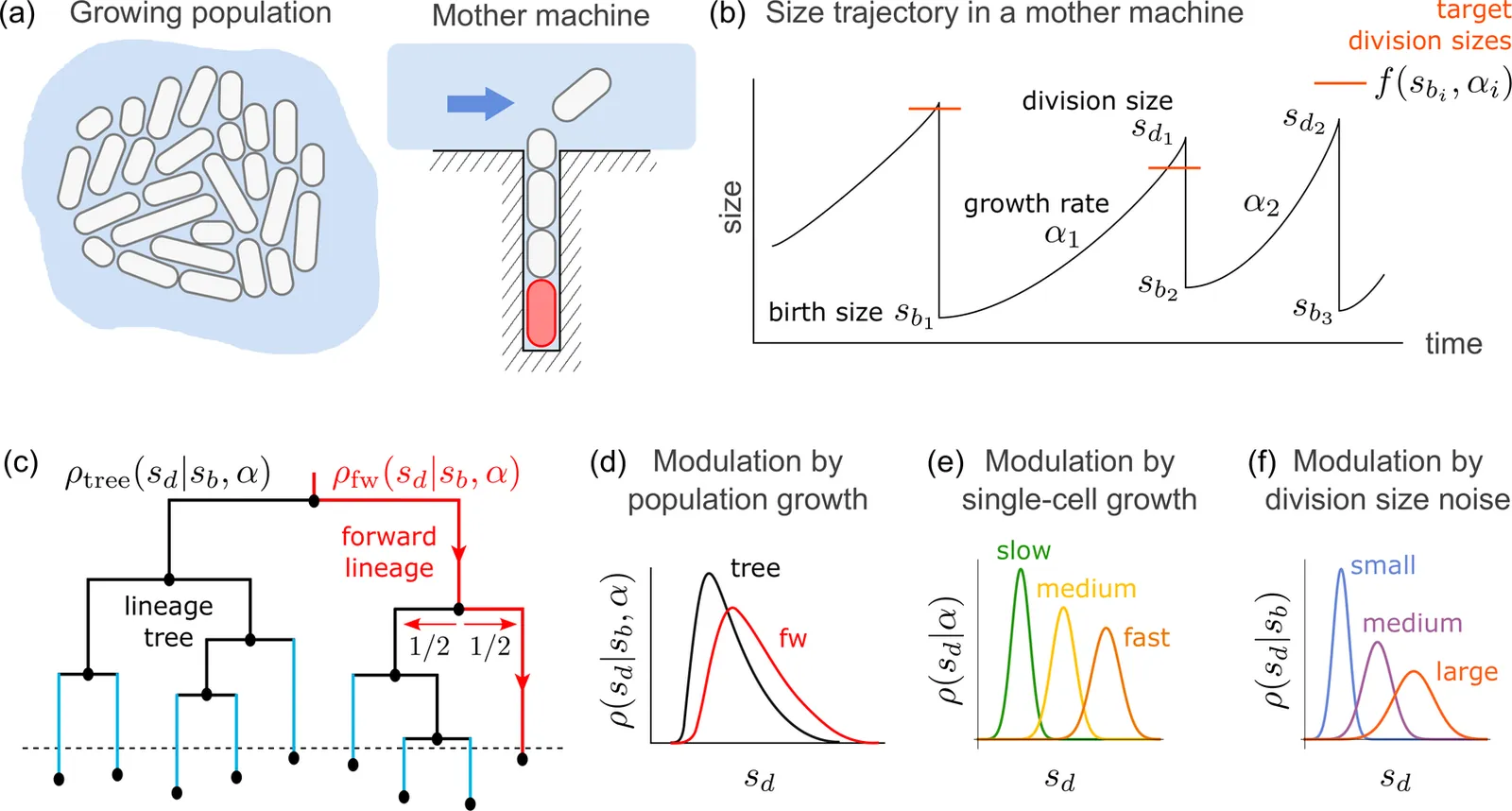
Living cells maintain size homeostasis by actively compensating for size fluctuations. Here, we present two stochastic maps that unify phenomenological models by integrating fluctuating single-cell growth rates and size-dependent noise mechanisms with cell size control. One map is applicable to mother machine lineages and the other to lineage trees of exponentially-growing cell populations, which reveals that population dynamics alter size control measured in mother machine experiments. For example, an adder can become more sizer-like or more timer-like at the population level depending on the noise statistics. Our analysis of bacterial data identifies extrinsic noise as the dominant mechanism of size variability, characterized by a quadratic conditional variance-mean relationship for division size across growth conditions. This finding contradicts the reported independence of added size relative to birth size but is consistent with the adder property in terms of the independence of the mean added size. Finally, we derive a trade-off between population-growth-rate gain and division-size noise. Correlations between size control quantifiers and single-cell growth rates inferred from data indicate that bacteria prioritize a narrow division-size distribution over growth rate maximisation.
We explore the connection between evolution and large-deviation theory. To do so, we study evolutionary dynamics in which individuals experience mutations, reproduction, and selection using variants of the Moran model. We show that, in the large population size limit, the impact of reproduction and selection amounts to realizing a large-deviation dynamics for the non-interacting random walk in which individuals simply explore the genome landscape due to mutations. This mapping, which holds at all times, allows us to recast transitions in the population genome distribution as dynamical phase transitions, which can then be studied using the toolbox of large-deviation theory. Finally, we show that the mapping extends beyond the class of Moran models.
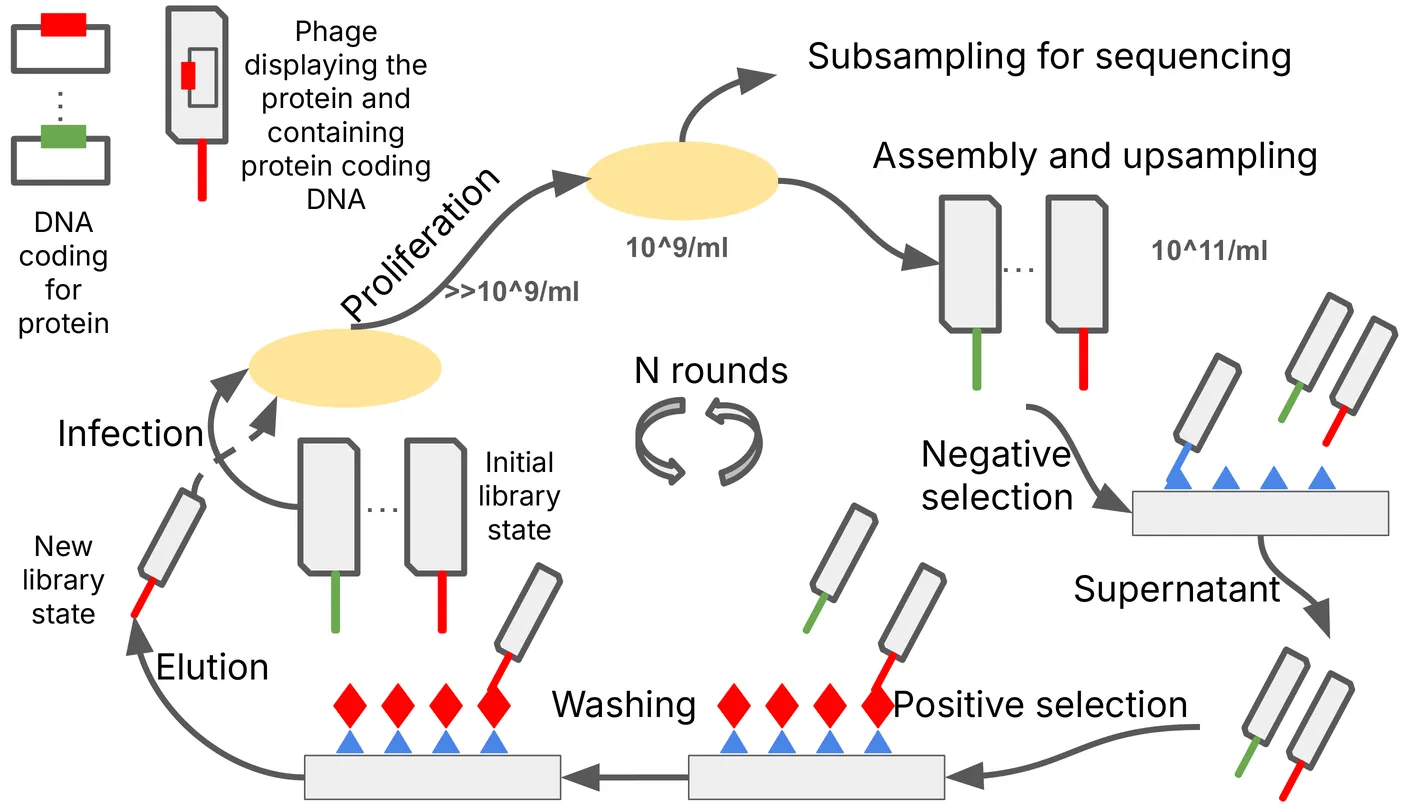
Phage display is a powerful laboratory technique used to study the interactions between proteins and other molecules, whether other proteins, peptides, DNA or RNA. The under-utilisation of this data in conjunction with deep learning models for protein design may be attributed to; high experimental noise levels; the complex nature of data pre-processing; and difficulty interpreting these experimental results. In this work, we propose a novel approach utilising a Bayesian Neural Network within a training loop, in order to simulate the phage display experiment and its associated noise. Our goal is to investigate how understanding the experimental noise and model uncertainty can enable the reliable application of such models to reliably interpret phage display experiments. We validate our approach using actual binding affinity measurements instead of relying solely on proxy values derived from 'held-out' phage display rounds.
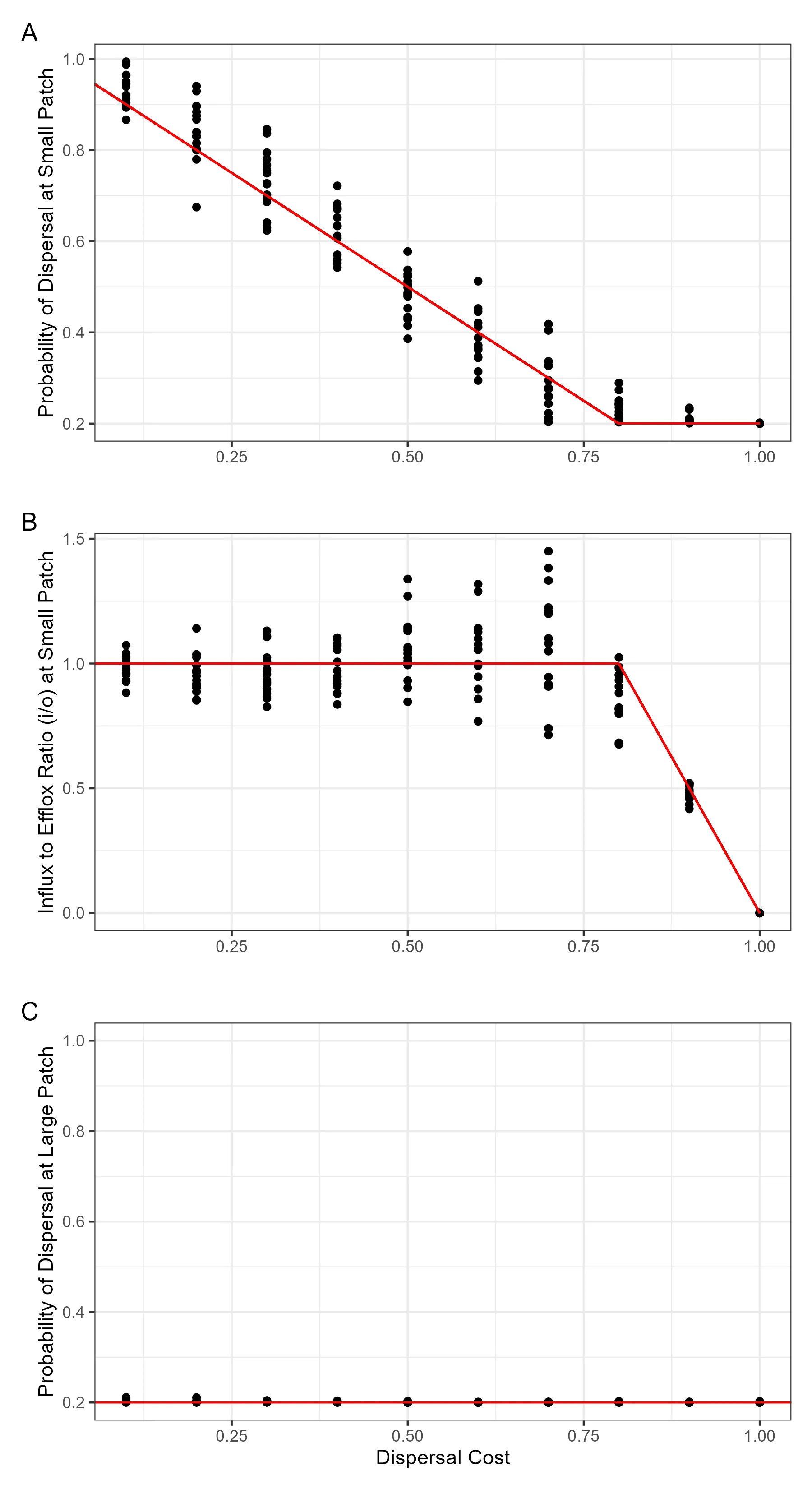
Fitness consequence of dispersal depends on property of the entire landscape, which patches are available and what are the cost of moving. These are information that are not available locally when an organism make the decision to disperse. This poses a problem to the organism, where it is unclear how an adaptive decision can be made. This also poses a problem to the scientist, since in order to study the adaptiveness of dispersal, we need information of the entire landscape. For theorist, this is through making a series of assumption about either the landscape or the organism, and for empiricists, this means a large amount of measurements needs to be made across a large area. In this paper, we propose a link between local demographic process, which an organism can have access to, to the fitness consequence of dispersal. This meant local environmental cue can be used for the decision on dispersal, and hence allow the evolution of plastic dispersal strategy. We will then show that using this approach, evolution of dispersal on complex landscape can be modelled with relative ease, and to show that accidental dispersal in one patch can drive the evolution of adaptive dispersal in another.
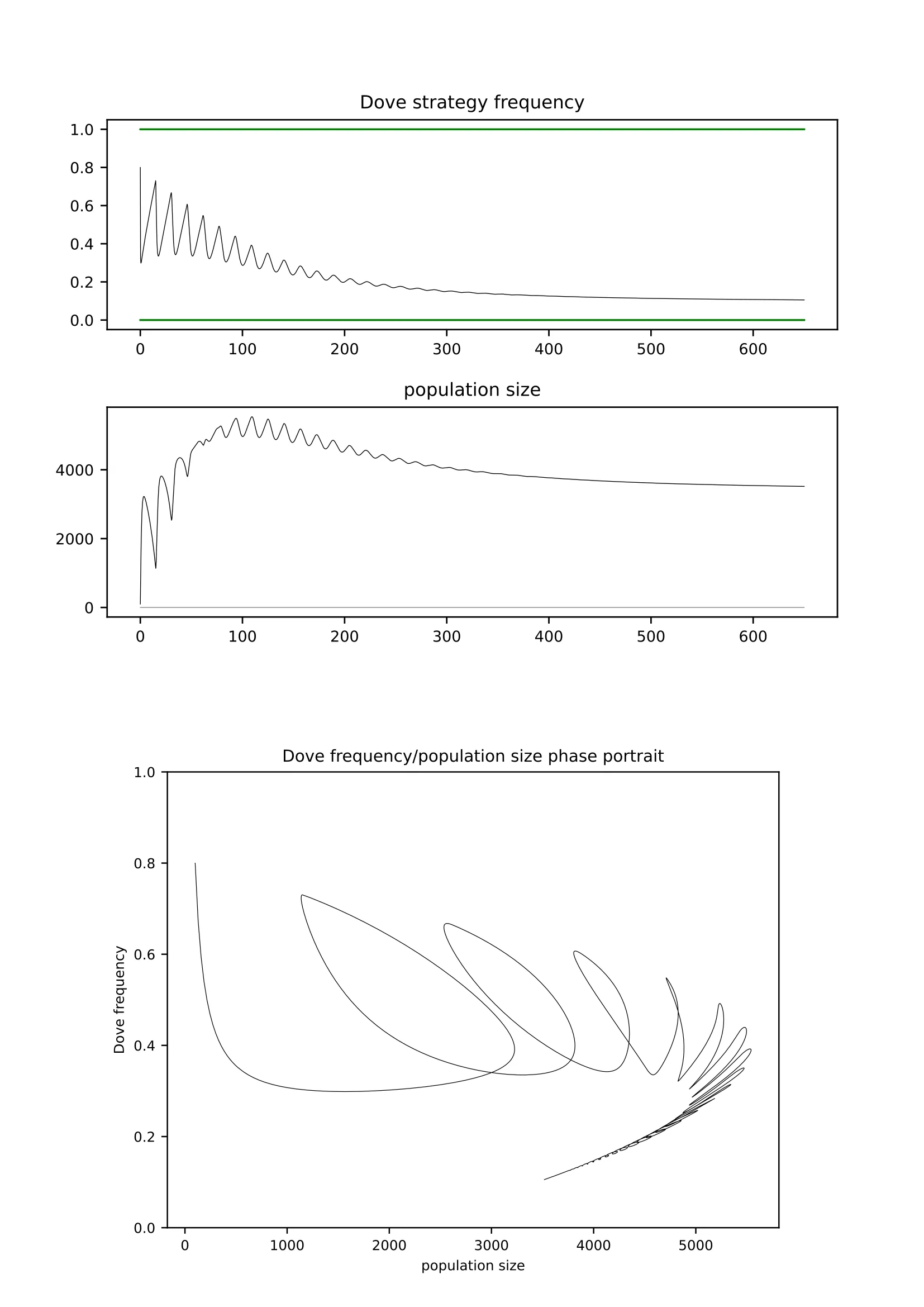
In this paper, we extend the demographic eco-evolutionary game approach, based on explicit birth and death dynamics instead of abstract "fitness" interpreted as an abstract "Malthusian parameter", by the introduction of the delay resulting from the juvenile maturation time. This leads to the application of the Delay Differential Equations (DDE). We show that delay seriously affects the resulting dynamics and may lead to the loss of stability of equilibria when critical delay is exceeded. We provide theoretical tools for the assessment of the critical delays and the parameter values when this may happen. Our results emphasize the importance of the mechanisms of density dependence. We analyze the impact of three different suppression modes based on: adult mortality, juvenile recruitment survival after the maturation period (without delay), and juvenile recruitment at birth (with the delay). The last mode leads to extreme patterns such as bifurcations, complex cycles, and chaotic dynamics. However, surprisingly, this mode leads to extension of the duration of the temporary transient metastable states known as "ghost attractors". In addition, we also focus on the problem of resilience of the analyzed systems against external periodic perturbations and feedback-driven factors such as additional predator pressure.
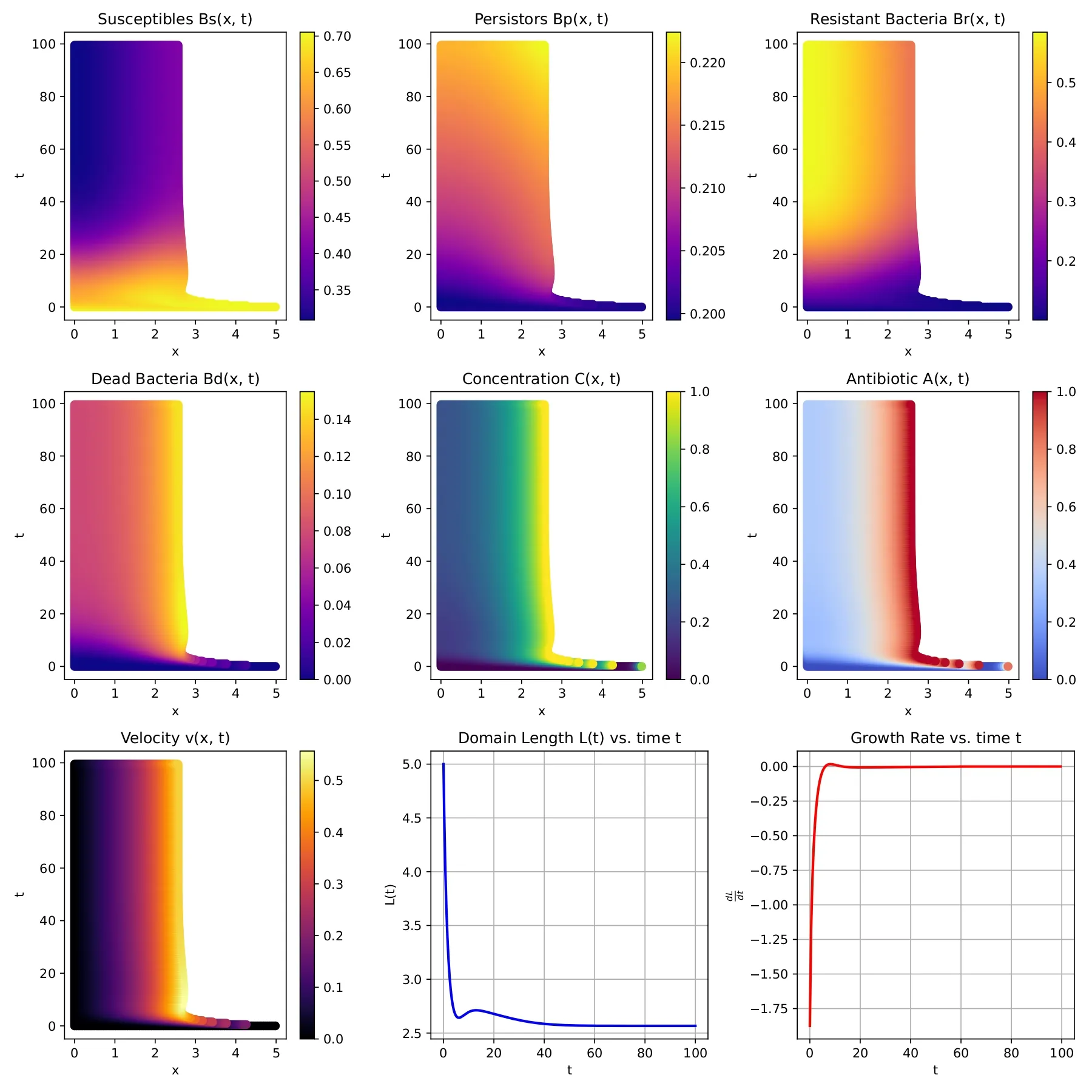
We propose and analyze a model for antibiotic resistance transfer in a bacterial biofilm and examine antibiotic dosing strategies that are effective in bacterial elimination. In particular, we consider a 1-D model of a biofilm with susceptible, persistor and resistant bacteria. Resistance can be transferred to the susceptible bacteria via horizontal gene transfer (HGT), specifically via conjugation. We analyze some basic properties of the model, determine the conditions for existence of disinfection and coexistence states, including boundary equilibria and their stability. Numerical simulations are performed to explore different modeling scenarios and support our theoretical findings. Different antibiotic dosing strategies are then studied, starting with a continuous dosing; here we note that high doses of antibiotic are needed for bacterial elimination. We then consider periodic dosing, and again observe that insufficient levels of antibiotic per dose may lead to treatment failure. Finally, using an extended version of Pontryagin's maximum principle we determine efficient antibiotic dosing protocols, which ensure bacterial elimination while keeping the total dosing low; we note that this involves a tapered dosing which reinforces results presented in other clinical and modeling studies. We study the optimal dosing for different parameter values and note that the optimal dosing schedule is qualitatively robust.

We investigate a type of lunar calendar known as lists of the 'nights of the moon', found throughout East Polynesia, including Rapa Nui (Easter Island). Using computational methods, we analyzed the lexical and structural divergence of 49 calendric lists from all major archipelagos, each containing about 30 night names. Our results, presented as a rooted phylogenetic tree, show a clear split into two main groups: one including lists from Rapa Nui, Mangareva, and the Marquesas; the other comprising lists from New Zealand, Hawaii, the Cook Islands, the Austral Islands, Tahiti, and the Tuamotu. This pattern aligns with a recent alternative classification of East Polynesian languages into 'Distal' (Marquesan, Mangarevan, Rapanui) and 'Proximal' (Maori, Hawaiian, Tahitian, etc.) subgroups. Since both language and lunar calendars are symbolic systems passed down and changed within communities - and given the geographic isolation of many archipelagos - we interpret this correspondence as evidence that the early divergence of East Polynesian lunar calendars mirrors early population movements and language splits in the region.
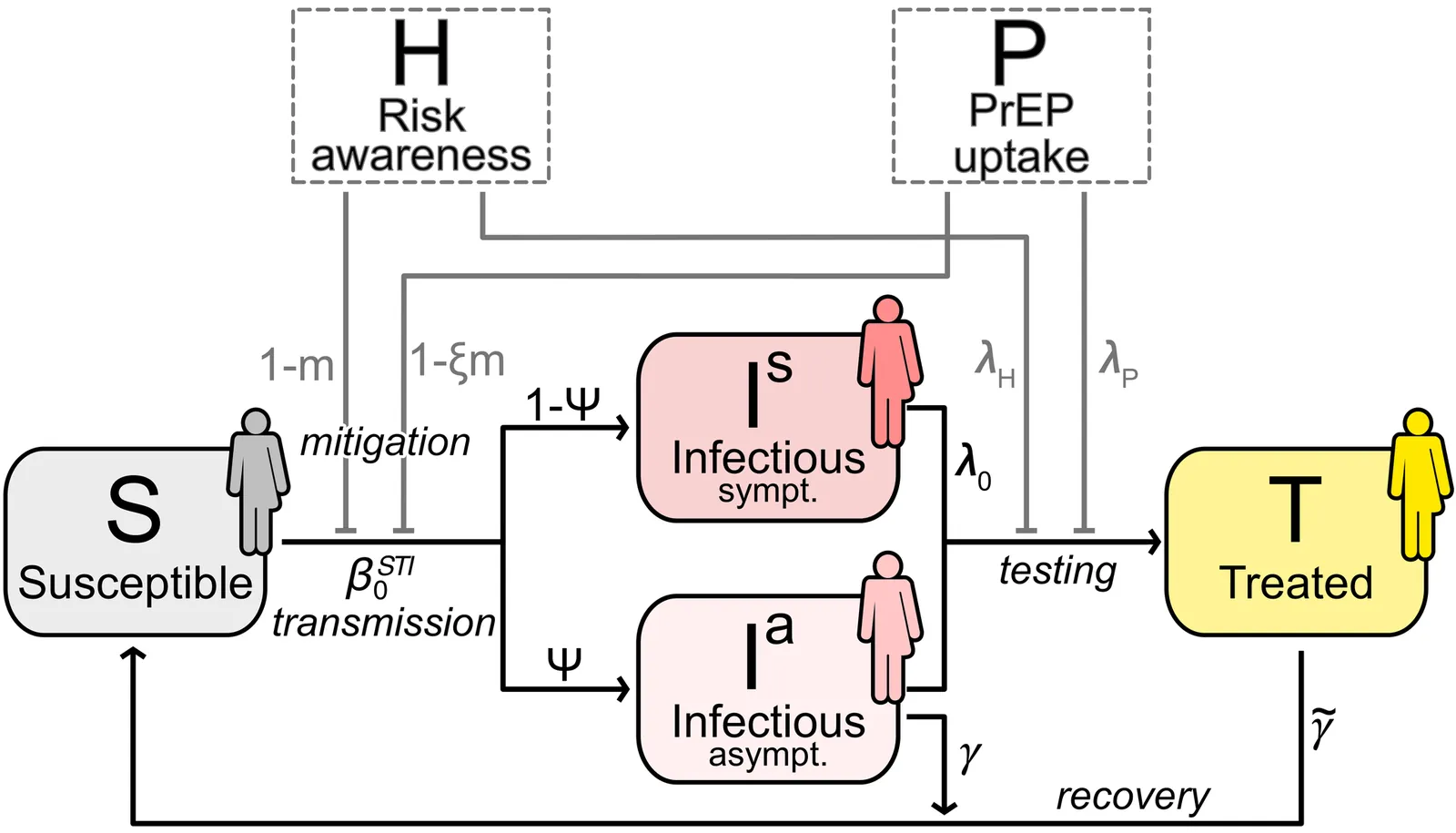
HIV pre-exposure prophylaxis (PrEP) drastically reduces the risk of HIV infection if taken as prescribed, providing almost perfect protection even during unprotected sexual intercourse. Although this has been transformative in reducing new HIV infections among high-risk populations, it has also been linked to an increase in risk practices -- a phenomenon known as risk compensation -- thereby favoring the spread of other sexually transmitted infections (STIs) deemed less severe. In this paper, we study a minimal compartmental model describing the effect of risk awareness and risk compensation due to PrEP on the spread of other STIs among a high-infection-risk group of men who have sex with men (MSM). The model integrates three key elements of risk-mediated behavior and PrEP programs: (i) HIV risk awareness drives self-protective behaviors (such as condom use and voluntary STI screening); (ii) individuals on PrEP are subject to risk compensation, but (iii) are required to screen for asymptomatic STIs frequently. We derived the basic reproduction number of the system, $R_0$, and found a transcritical bifurcation at $R_0=1$, where the disease-free equilibrium becomes unstable and an endemic equilibrium emerges. This endemic equilibrium is asymptotically stable wherever it exists. We identified critical thresholds in behavioral and policy parameters that separate these regimes and analyzed typical values for plausible parameter choices. Beyond the specific epidemiological context, the model serves as a general framework for studying nonlinear interactions between behavioral adaptation, preventive interventions, and disease dynamics, providing insights into how feedback mechanisms can lead to non-trivial responses in epidemic systems. Finally, our model can be easily extended to study the effect of interventions and risk compensation in other STIs.
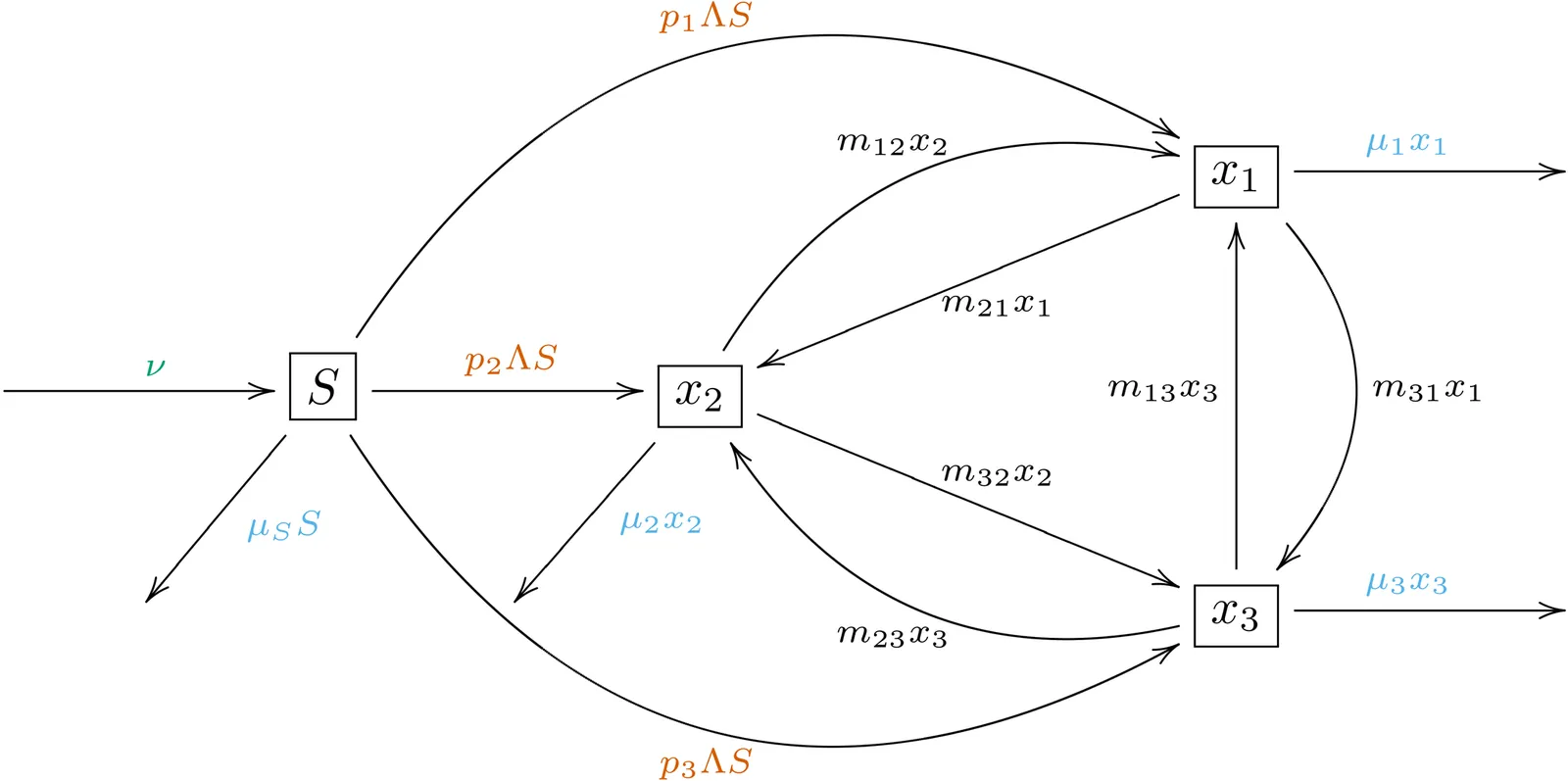
Transmission dynamics of infectious diseases are often studied using compartmental mathematical models, which are commonly represented as systems of autonomous ordinary differential equations. A key step in the analysis of such models is to identify equilibria and find conditions for their stability. Local stability analysis reduces to a problem in linear algebra, but there is no general algorithm for establishing global stability properties. Substantial progress on global stability of epidemic models has been made in the last 20 years, primarily by successfully applying Lyapunov's method to specific systems. Here, we show that any compartmental epidemic model in which susceptible individuals cannot be distinguished and can be infected only once, has a globally asymptotically stable (GAS) equilibrium. If the basic reproduction number ${R}_0$ satisfies ${R}_0 > 1$, then the GAS fixed point is an endemic equilibrium (i.e., constant, positive disease prevalence). Alternatively, if ${R}_0 \le 1$, then the GAS equilibrium is disease-free. This theorem subsumes a large number of results published over the last century, strengthens most of them by establishing global rather than local stability, avoids the need for any stability analyses of these systems in the future, and settles the question of whether co-existing stable solutions or non-equilibrium attractors are possible in such models: they are not.
Community science observational datasets are useful in epidemiology and ecology for modeling species distributions, but the heterogeneous nature of the data presents significant challenges for standardization, data quality assurance and control, and workflow management. In this paper, we present a data workflow for cleaning and harmonizing multiple community science datasets, which we implement in a case study using eBird, iNaturalist, GBIF, and other datasets to model the impact of highly pathogenic avian influenza in populations of birds in the subantarctic. We predict population sizes for several species where the demographics are not known, and we present novel estimates for potential mortality rates from HPAI for those species, based on a novel aggregated dataset of mortality rates in the subantarctic.
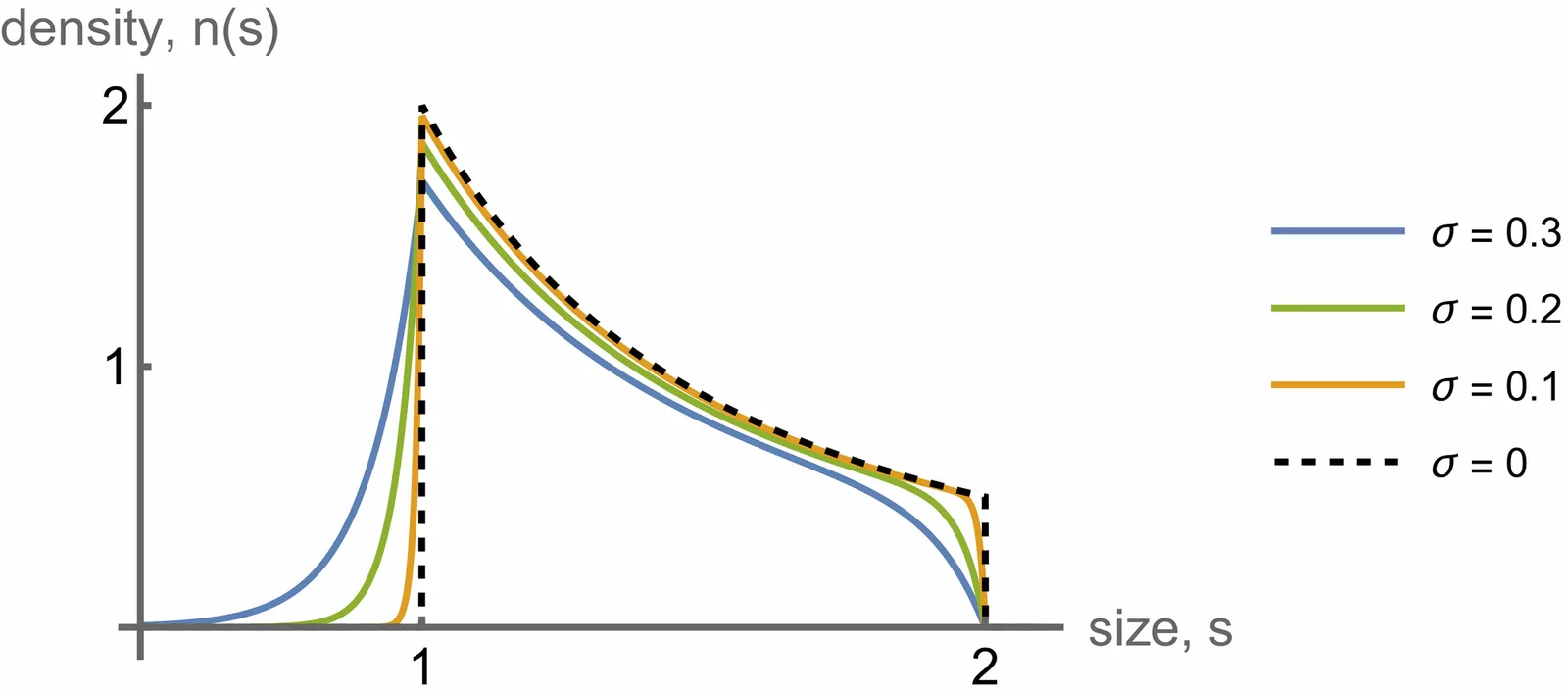
We analyze a size-structured branching process in which individual cells grow exponentially according to a Feller square-root process and divide under general size-control mechanisms. We obtain exact expressions for the asymptotic population growth rate, the steady-state snapshot distribution of cell sizes, and the fluctuations of the total cell number. Our first result is that the population growth rate is exactly equal to the mean single-cell growth rate, for all noise strengths and for all division and size-regulation schemes that maintain size homeostasis. Thus square-root growth noise is neutral with respect to long-term fitness, in sharp contrast to models with size-independent stochastic growth rates. Second, we show that the steady-state population cell-size distribution is obtained from the deterministic inverse-square-law solution by a one-sided exponential convolution with kernel width set by the strength of growth fluctuations. Third, the mean-rescaled population size $N_t/\left\langle N_t\right\rangle$ converges to a stationary compound Poisson-exponential distribution that depends only on growth noise. This distribution, and hence the long-time shape of population-size fluctuations, is unchanged by division-size noise or asymmetric partitioning. These results identify Feller-type exponential growth with square-root noise as an exactly solvable benchmark for stochastic growth in size-controlled populations and provide concrete signatures that distinguish it from models with size-independent growth-rate noise.

This study presents an agent-based model (ABM) developed to simulate staff and resident interactions within a synthetic aged care facility, capturing movement, task execution, and proximity-based contact events across three staff shifts and varying levels of resident care. Contacts were defined by spatial thresholds (1.5 m and 3 m) and cumulative duration, enabling the generation of detailed contact matrices. Simulation results showed that low and medium care residents experienced the highest frequency of interactions, particularly with staff on morning and afternoon shifts, while high care residents and night staff had substantially fewer contacts. Contact rates varied significantly by care level and shift, confirmed through Poisson-based regression modelling. Temporal analyses revealed clustering of high-risk contacts during structured daily routines, especially communal and care activities. An integrated airborne transmission module, seeded with a single infectious staff member, demonstrated that infection risk was highest during high-contact shifts and among medium care residents. Vaccination scenarios reduced predicted transmission by up to 68\%, with the greatest impact observed when both staff and residents were vaccinated. These findings highlight the importance of accounting for contact heterogeneity in aged care and demonstrate the utility of ABMs for evaluating targeted infection control strategies in high-risk, enclosed environments.

Large animal groups -- bird flocks, fish schools, insect swarms -- are often assumed to form by gradual aggregation of sparsely distributed individuals. Using a mathematically precise framework based on time-varying directed interaction networks, we show that this widely held view is incomplete. The theory demonstrates that large moving groups do not arise by slow accumulation; instead, they emerge through the rapid merging of multiple pre-existing subgroups that are simultaneously activated under high-density conditions. The key mechanism is topological: the long-term interaction structure of any moving group contains a single dominant strongly connected component (SCC). This dominant SCC determines the collective velocity -- both speed and direction -- of the entire group. When two subgroups encounter one another, the trailing subgroup aligns with -- and ultimately inherits -- the velocity of the dominant SCC of the leading subgroup. Repeated merging events naturally generate large groups whose speed is predicted to be lower than the mean speed of the original subgroups. The same dynamics explain several universal empirical features: broad neighbour-distance distributions, directional asymmetry in neighbour selection, and the characteristic narrow-front, wide-rear geometry of real flocks. The framework yields testable predictions for STARFLAG-style 3D datasets, offering a unified explanation for the formation, maintenance, and geometry of coordinated animal groups.
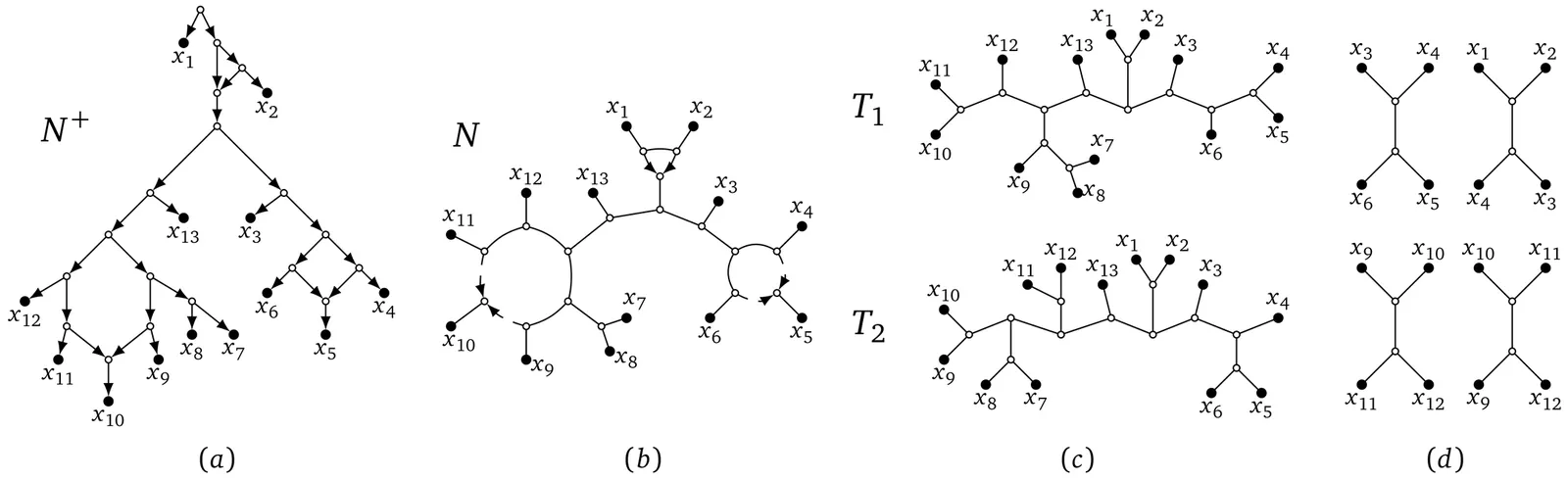
Phylogenetic trees and networks are graphs used to model evolutionary relationships, with trees representing strictly branching histories and networks allowing for events in which lineages merge, called reticulation events. While the question of data sufficiency has been studied extensively in the context of trees, it remains largely unexplored for networks. In this work we take a first step in this direction by establishing bounds on the amount of genomic data required to reconstruct binary level-$1$ semi-directed phylogenetic networks, which are binary networks in which reticulation events are indicated by directed edges, all other edges are undirected, and cycles are vertex-disjoint. For this class, methods have been developed recently that are statistically consistent. Roughly speaking, such methods are guaranteed to reconstruct the correct network assuming infinitely long genomic sequences. Here we consider the question whether networks from this class can be uniquely and correctly reconstructed from finite sequences. Specifically, we present an inference algorithm that takes as input genetic sequence data, and demonstrate that the sequence length sufficient to reconstruct the correct network with high probability, under the Cavender-Farris-Neyman model of evolution, scales logarithmically, polynomially, or polylogarithmically with the number of taxa, depending on the parameter regime. As part of our contribution, we also present novel inference rules for quartet data in the semi-directed phylogenetic network setting.

We considered a model for an infectious disease outbreak, when the depletion of susceptible individuals is negligible, and assumed that individuals adapt their behavior according to the information they receive about new cases. In line with the information index approach, we supposed that individuals react to past information according to a memory kernel that is continuously distributed in the past. We analyzed equilibria and their stability, with analytical results for selected cases. Thanks to the recently developed pseudospectral approximation of delay equations, we studied numerically the long-term dynamics of the model for memory kernels defined by gamma distributions with a general non-integer shape parameter, extending the analysis beyond what is allowed by the linear chain trick. In agreement with previous studies, we showed that behavior adaptation alone can cause sustained waves of infections even in an outbreak scenario, and notably in the absence of other processes like demographic turnover, seasonality, or waning immunity. Our analysis gives a more general insight into how the period and peak of epidemic waves depend on the shape of the memory kernel and how the level of minimal contact impacts the stability of the behavior-induced positive equilibrium.
We look at the interaction of dispersal and environmental stochasticity in $n$-patch models. We are able to prove persistence and extinction results even in the setting when the dispersal rates are stochastic. As applications we look at Beverton-Holt and Hassell functional responses. We find explicit approximations for the total population size at stationarity when we look at slow and fast dispersal. In particular, we show that if dispersal is small then in the Beverton-Holt setting, if the carrying capacity is random, then environmental fluctuations are always detrimental and decrease the total population size. Instead, in the Hassell setting, if the inverse of the carrying capacity is made random, then environmental fluctuations always increase the population size. Fast dispersal can save populations from extinction and therefore increase the total population size. We also analyze a different type of environmental fluctuation which comes from switching environmental states according to a Markov chain and find explicit approximations when the switching is either fast or slow - in examples we are able to show that slow switching leads to a higher population size than fast switching. Using and modifying some approximation results due to Cuello, we find expressions for the total population size in the $n=2$ patch setting when the growth rates, carrying capacities, and dispersal rates are influenced by random fluctuations. We find that there is a complicated interaction between the various terms and that the covariances between the various random parameters (growth rate, carrying capacity, dispersal rate) play a key role in whether we get an increase or a decrease in the total population size. Environmental fluctuations turn to sometimes be beneficial -- this show that not only dispersal, but also environmental stochasticity can lead to an increase in population size.
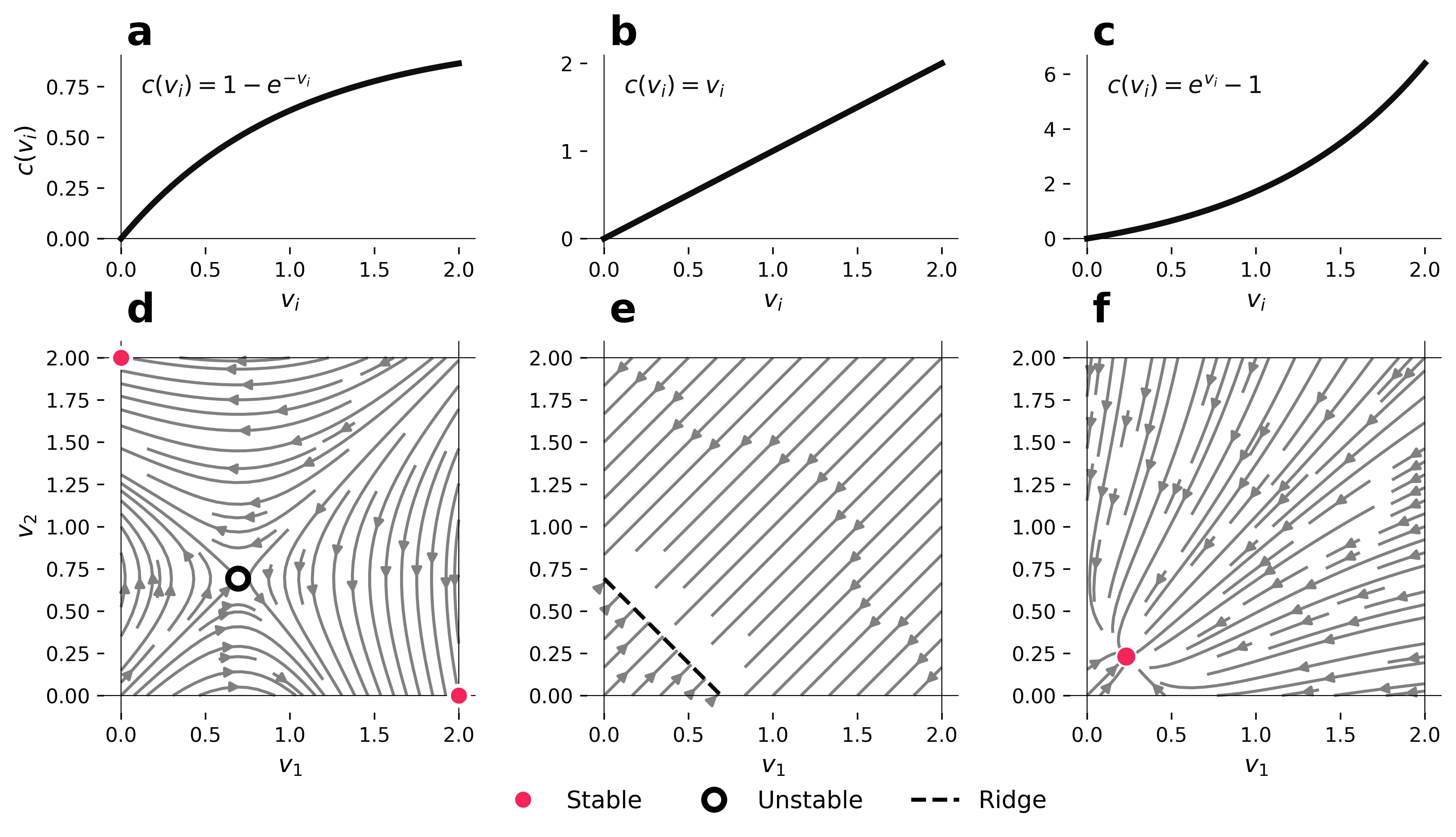
Collective vigilance describes how animals in groups benefit from the predator detection efforts of others. Empirical observations typically find either a many-eyes strategy with all (or many) group members maintaining a low level of individual vigilance, or a sentinel strategy with one (or a few) individuals maintaining a high level of individual vigilance while others do not. With a general analytical treatment that makes minimal assumptions, we show that these two strategies are alternate solutions to the same adaptive problem of balancing the costs of predation and vigilance. Which strategy is preferred depends on how costs scale with the level of individual vigilance: many-eyes strategies are preferred where costs of vigilance rise gently at low levels but become steeper at higher levels (convex; e.g. an open field); sentinel strategies are preferred where costs of vigilance rise steeply at low levels and then flatten out (concave; e.g. environments with vantage points). This same dichotomy emerges whether individuals act selfishly to optimise their own fitness or cooperatively to optimise group fitness. The model is extended to explain discrete behavioural switching between strategies and differential levels of vigilance such as edge effects.
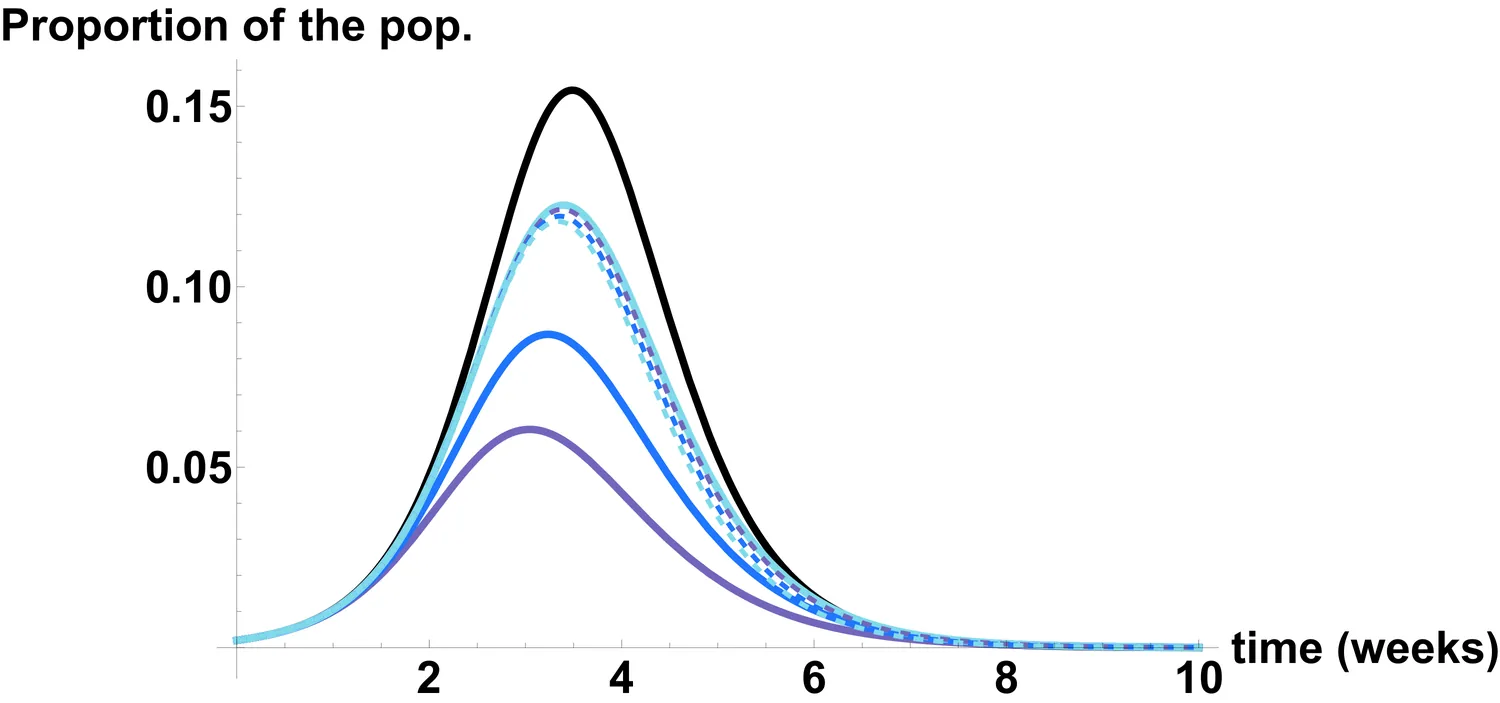
Population heterogeneity is a key factor in epidemic dynamics, influencing both transmission and final epidemic size. While heterogeneity is often modeled through age structure, spatial location, or contact patterns, differences in host susceptibility have recently gained attention, particularly during the COVID-19 pandemic. Building on the framework of Diekmann and Inaba (Journal of Mathematical Biology, 2023), we focus on the special case of SEIR-models, which are widely used for influenza and other respiratory infections. We derive the model equations under two distinct assumptions linking susceptibility and infectiousness. Analytical results show that heterogeneity in susceptibility reduces the epidemic final size compared to homogeneous models with the same basic reproduction number $\Ro$. In the case of gamma-distributed susceptibility, we obtain stronger results on the epidemic final size. The resulting model captures population heterogeneity through a single parameter, which makes it practical for fitting epidemic data. We illustrate its use by applying it to seasonal influenza in Italy.
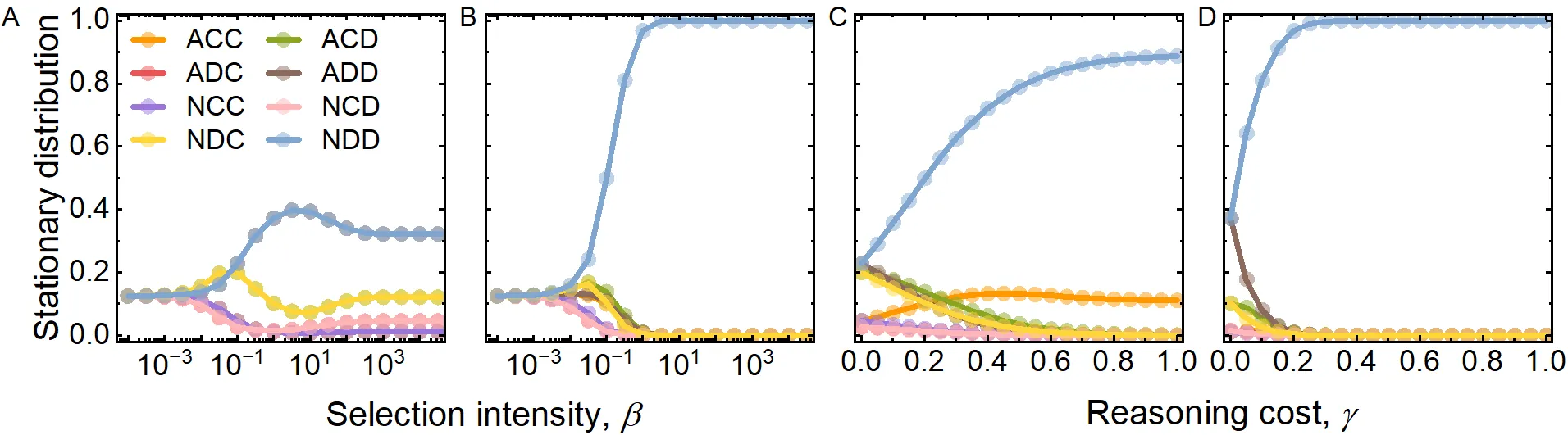
Understanding how cooperation emerges and persists is a central challenge in evolutionary game theory. Existing models often rely on restricted, hand-picked strategy sets, which can overlook critical behavioural pathways. A recent four-strategy framework showed that cheap talk can promote cooperation through local interactions, yet it remained unclear whether modelled strategies might alter these conclusions. Here, we extend this framework to the complete set of eight strategies that naturally arise from communication and decision-making rules. We show that incorporating the full strategy space dramatically changes the evolutionary landscape. Cooperation becomes both more robust and more versatile, driven by novel pathways absent in the restricted model. In particular, we uncover a previously overlooked mechanism in which suspicious cooperation catalyses a cyclic dynamic that sustains cooperation. Conversely, the assumed role of strategic defection in the biased model is fragile, acting mainly as a spoiler rather than a genuine evolutionary attractor. The complete model further reveals a rich spectrum of long-term behaviours, including stable coexistence among up to seven strategies and time-varying patterns of partial coexistence. These results demonstrate that the full strategy space unlocks hidden routes to cooperative behaviour and highlight the importance of comprehensive modelling when explaining the emergence of cooperation.

We study a model for a mosquito-borne epidemic outbreak in which humans can adopt protective behaviour against vector bites depending on information on the past and present prevalence. Assuming that mosquitoes can also feed on other non-competent hosts (i.e. hosts that cannot infect others), we first review some results from the literature by showing that protective behaviour may either decrease or increase the value of the reproduction number of the epidemic depending on multiple elements. Then, assuming that changes in opinion occur much faster than the spread of the disease, we exploit an approach based on the Geometric Singular Perturbation Theory to reduce the two-group model to a model for a homogeneous host population. Then, we use the resulting model to investigate the effect of information-induced behavioural changes on the transient dynamics of the epidemic, discussing the case when protective measures induced an outbreak with a low attack rate. We illustrate how behavioural changes might either help in containing an epidemic outbreak or make the epidemic last longer, even triggering recurrent damped epidemic waves. We conclude with numerical simulations to illustrate our analytical results.
Transmission dynamics of infectious diseases are often studied using compartmental mathematical models, which are commonly represented as systems of autonomous ordinary differential equations. A key step in the analysis of such models is to identify equilibria and find conditions for their stability. Local stability analysis reduces to a problem in linear algebra, but there is no general algorithm for establishing global stability properties. Substantial progress on global stability of epidemic models has been made in the last 20 years, primarily by successfully applying Lyapunov's method to specific systems. Here, we show that any compartmental epidemic model in which susceptible individuals cannot be distinguished and can be infected only once, has a globally asymptotically stable (GAS) equilibrium. If the basic reproduction number ${R}_0$ satisfies ${R}_0 > 1$, then the GAS fixed point is an endemic equilibrium (i.e., constant, positive disease prevalence). Alternatively, if ${R}_0 \le 1$, then the GAS equilibrium is disease-free. This theorem subsumes a large number of results published over the last century, strengthens most of them by establishing global rather than local stability, avoids the need for any stability analyses of these systems in the future, and settles the question of whether co-existing stable solutions or non-equilibrium attractors are possible in such models: they are not.
We considered a model for an infectious disease outbreak, when the depletion of susceptible individuals is negligible, and assumed that individuals adapt their behavior according to the information they receive about new cases. In line with the information index approach, we supposed that individuals react to past information according to a memory kernel that is continuously distributed in the past. We analyzed equilibria and their stability, with analytical results for selected cases. Thanks to the recently developed pseudospectral approximation of delay equations, we studied numerically the long-term dynamics of the model for memory kernels defined by gamma distributions with a general non-integer shape parameter, extending the analysis beyond what is allowed by the linear chain trick. In agreement with previous studies, we showed that behavior adaptation alone can cause sustained waves of infections even in an outbreak scenario, and notably in the absence of other processes like demographic turnover, seasonality, or waning immunity. Our analysis gives a more general insight into how the period and peak of epidemic waves depend on the shape of the memory kernel and how the level of minimal contact impacts the stability of the behavior-induced positive equilibrium.
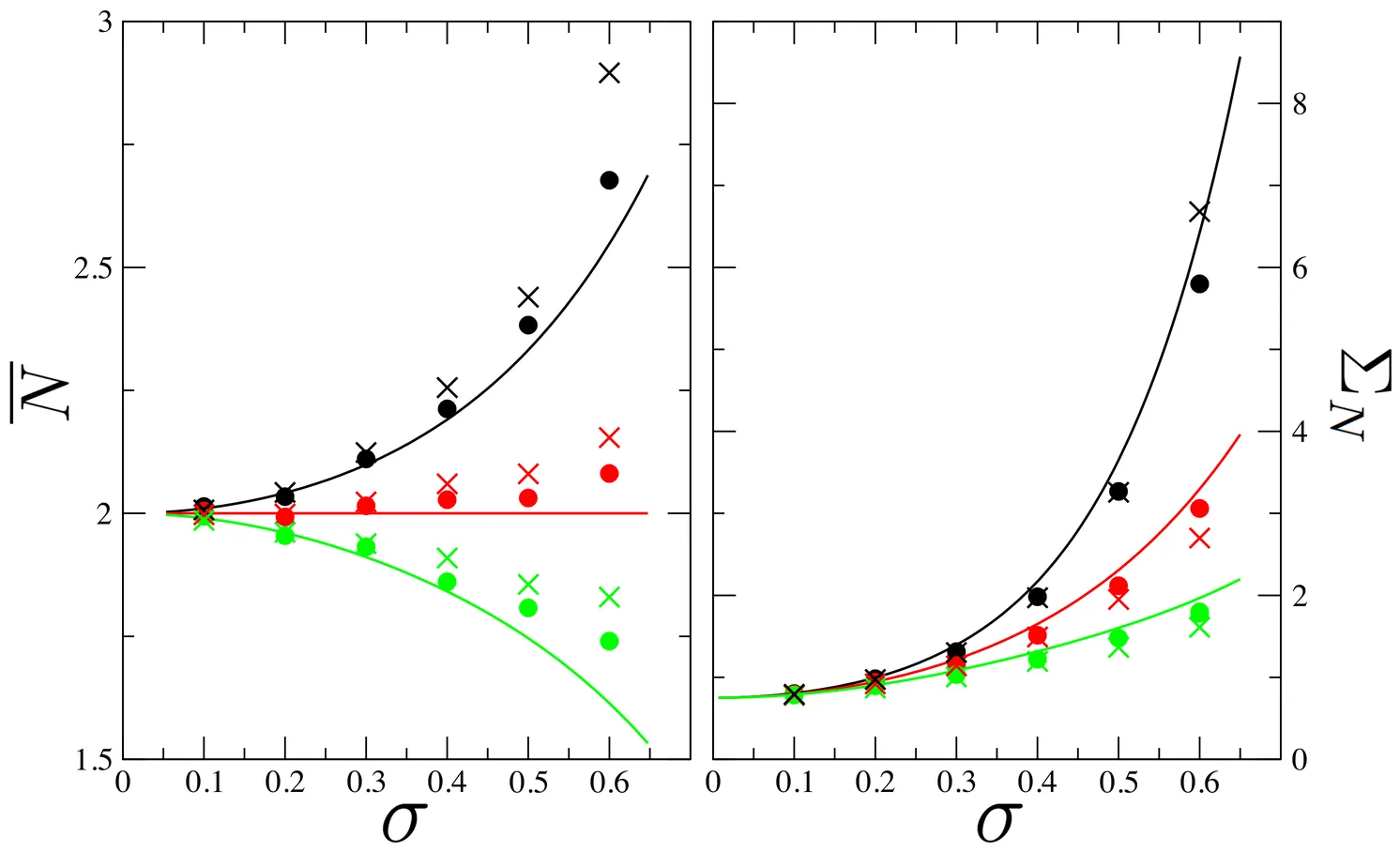
Multispecies ecosystems modelled by generalized Lotka-Volterra equations exhibit stationary population abundances, where large number of species often coexist. Understanding the precise conditions under which this is at all feasible and what triggers species extinctions is a key, outstanding problem in theoretical ecology. Using standard methods of random matrix theory, I show that distributions of species abundances are Gaussian at equilibrium, in the weakly interacting regime. One consequence is that feasibility is generically broken before stability, for large enough number of species. I further derive an analytic expression for the probability that $n=0,1,2,...$ species go extinct and conjecture that a single-parameter scaling law governs species extinctions. These results are corroborated by numerical simulations in a wide range of system parameters.
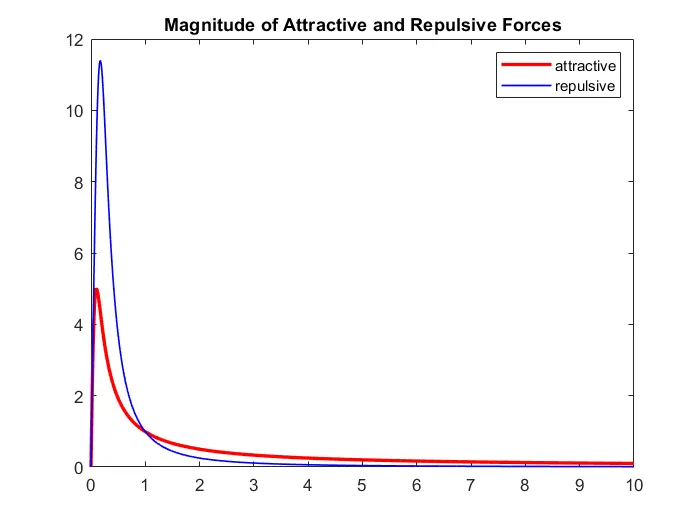
We investigate predator-prey school interactions in aquatic environments using a stochastic differential equation (SDE)-based, particle-level model that incorporates attraction, repulsion, alignment, and environmental noise. Two predation strategies-center attack and nearest attack-are examined to assess their effects on prey survival, predator efficiency, and group dynamics. Simulations reveal diverse emergent behaviors such as prey dispersal and regrouping, oscillatory predation with collective defense, and predator encirclement. Results show that collective hunting enhances capture efficiency compared to solitary attacks, but benefits diminish beyond a critical predator group size due to intra-predator competition. This work provides new insights into cooperative predation and introduces a generalizable SDE framework for analyzing predator-prey interactions.

Phylogenetic trees represent certain species and their likely ancestors. In such a tree, present-day species are leaves and an edge from u to v indicates that u is an ancestor of v. Weights on these edges indicate the phylogenetic distance. The phylogenetic diversity (PD) of a set of species A is the total weight of edges that are on any path between the root of the phylogenetic tree and a species in A. Selecting a small set of species that maximizes phylogenetic diversity for a given phylogenetic tree is an essential task in preservation planning, where limited resources naturally prevent saving all species. An optimal solution can be found with a greedy algorithm [Steel, Systematic Biology, 2005; Pardi and Goldman, PLoS Genetics, 2005]. However, when a food web representing predator-prey relationships is given, finding a set of species that optimizes phylogenetic diversity subject to the condition that each saved species should be able to find food among the preserved species is NP-hard [Spillner et al., IEEE/ACM, 2008]. We present a generalization of this problem, where, inspired by biological considerations, the food web has weighted edges to represent the importance of predator-prey relationships. We show that this version is NP-hard even when both structures, the food web and the phylogenetic tree, are stars. To cope with this intractability, we proceed in two directions. Firstly, we study special cases where a species can only survive if a given fraction of its prey is preserved. Secondly, we analyze these problems through the lens of parameterized complexity. Our results include that finding a solution is fixed-parameter tractable with respect to the vertex cover number of the food web, assuming the phylogenetic tree is a star.
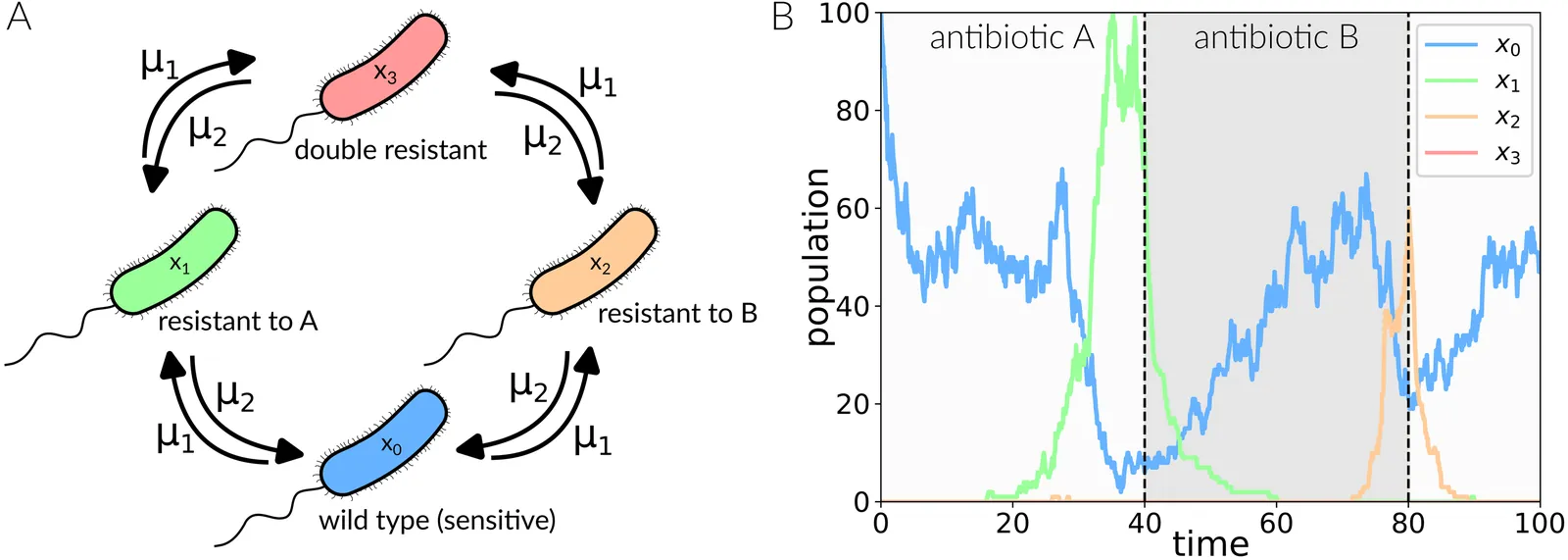
Antimicrobial resistance (AMR) threatens global health. A promising and underexplored strategy to tackle this problem are sequential therapies exploiting collateral sensitivity (CS), whereby resistance to one drug increases sensitivity to another. Here, we develop a four-genotype stochastic birth-death model with two bacteriostatic antibiotics to identify switching periods that maximize bacterial extinction under subinhibitory concentrations. We show that extinction probability depends nonlinearly on switching period, with stepwise increases aligned to discrete switch events: fast sequential therapies are suboptimal as they do not allow for the evolution of resistance, a key ingredient in these therapies. A geometric distribution framework accurately predicts cumulative extinction probabilities, where the per-switch extinction probability rises with switching period. We further derive a heuristic approximation for the extinction probability based on times to fixation of single-resistant mutants. Sensitivity analyses reveal that strong reciprocal CS is required for this strategy to work, and we explore how increasing antibiotic doses and higher mutation rates modulate extinction in a nonmonotonic manner. Finally, we discuss how longer therapies maximize extinction but also cause higher resistance, leading to a Pareto front of optimal switching periods. Our results provide quantitative design principles for in vitro and clinical sequential antibiotic therapies, underscoring the potential of CS-guided regimens to suppress resistance evolution and eradicate infections.

Explaining the emergence of self-organized biodiversity and species abundance distribution patterns remians a fundamental challenge in ecology. While classical frameworks, such as neutral theory and models based on pairwise species interactions, have provided valuable insights, they often neglect higher-order interactions (HOIs), whose role in stabilizing ecological communities is increasingly recognized. Here, we extend the Generalized Lotka-Volterra framework to incorporate HOIs and demonstrate that these interactions can enhance ecosystem stability and prevent collapse. Our model exhibits a diverse range of emergent dynamics, including self-sustained oscillations, quasi-periodic (torus) trajectories, and intermittent chaos. Remarkably, it also reproduces empirical species abundance distributions observed across diverse natural communities. These results underscore the critical role of HOIs in structuring biodiversity and offer a broadly applicable theoretical framework for capturing complexity in ecological systems
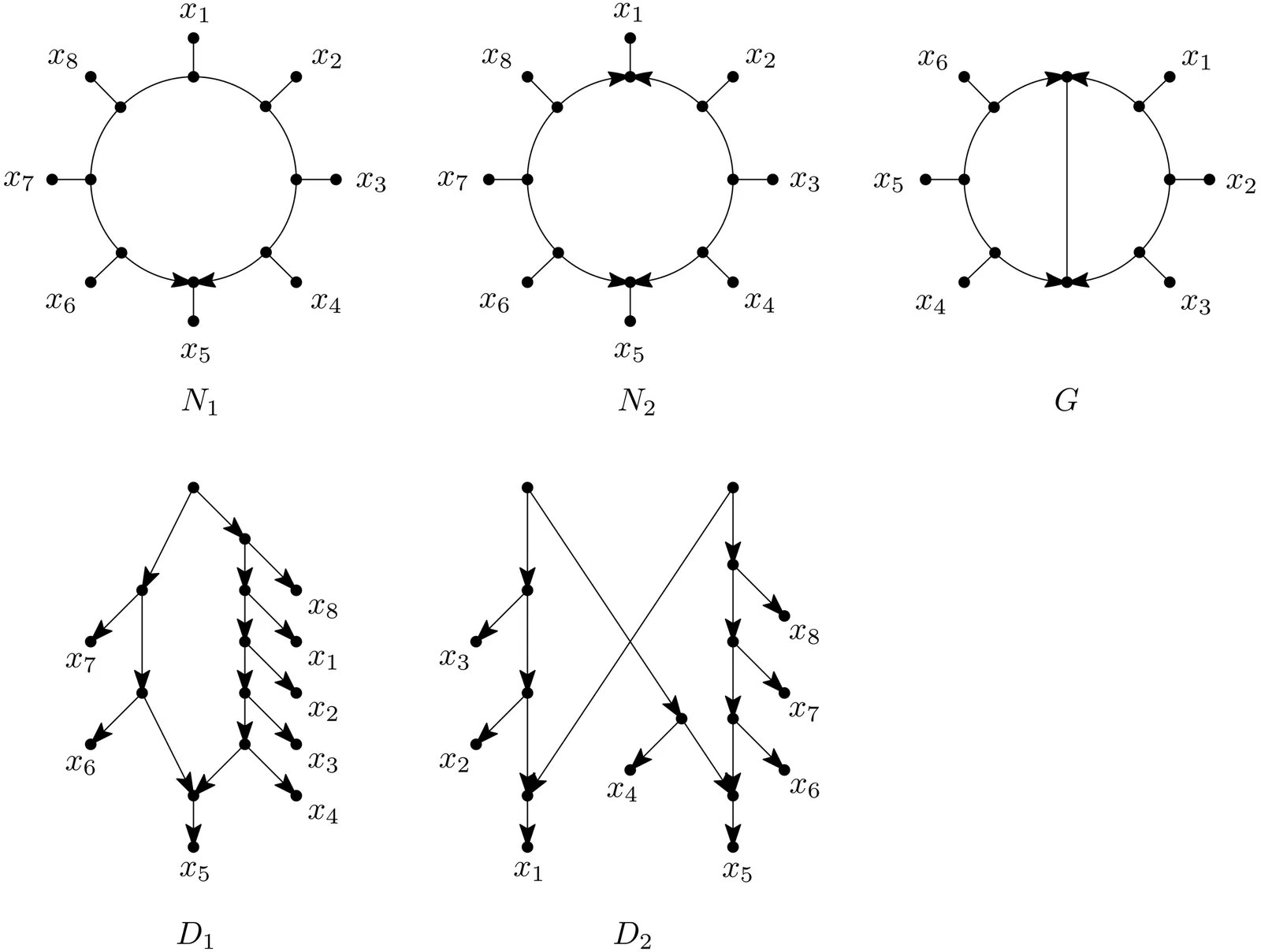
In evolutionary biology, phylogenetic networks are graphs that provide a flexible framework for representing complex evolutionary histories that involve reticulate evolutionary events. Recently phylogenetic studies have started to focus on a special class of such networks called semi-directed networks. These graphs are defined as mixed graphs that can be obtained by de-orienting some of the arcs in some rooted phylogenetic network, that is, a directed acyclic graph whose leaves correspond to a collection of species and that has a single source or root vertex. However, this definition of semi-directed networks is implicit in nature since it is not clear when a mixed-graph enjoys this property or not. In this paper, we introduce novel, explicit mathematical characterizations of semi-directed networks, and also multi-semi-directed networks, that is, mixed graphs that can be obtained from directed phylogenetic networks that may have more than one root. In addition, through extending foundational tools from the theory of rooted networks into the semi-directed setting - such as cherry picking sequences, omnians, and path partitions - we characterize when a (multi-)semi-directed network can be obtained by de-orienting some rooted network that is contained in one of the well-known classes of tree-child, orchard, tree-based or forest-based networks. These results address structural aspects of (multi-)semi-directed networks and pave the way to improved theoretical and computational analyses of such networks, for example, within the development of algebraic evolutionary models that are based on such networks.

Decades of scientific inquiry have sought to understand how evolution fosters cooperation, a concept seemingly at odds with the belief that evolution should produce rational, self-interested individuals. Most previous work has focused on the evolution of cooperation among boundedly rational individuals whose decisions are governed by behavioral rules that do not need to be rational. Here, using an evolutionary model, we study how altruism can evolve in a community of rational agents and promote cooperation. We show that in both well-mixed and structured populations, a population of objectively rational agents is readily invaded by mutant individuals who make rational decisions but evolve a distorted (i.e., subjective) perception of their payoffs. This promotes behavioral diversity and gives rise to the evolution of rational, other-regarding agents who naturally solve all the known strategic problems of two-person, two-strategy games by perceiving their games as pure coordination games.
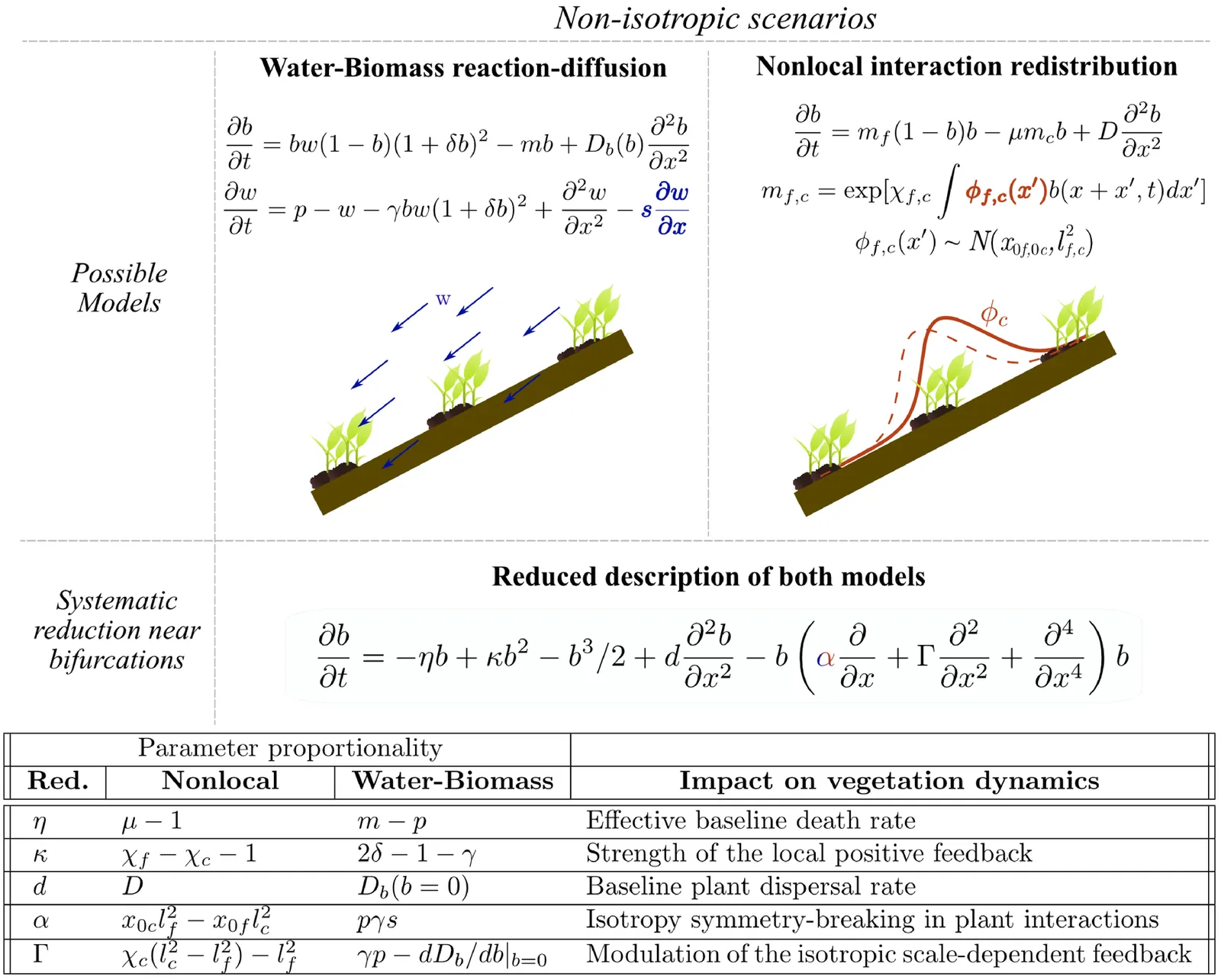
Several theoretical models predict that spatial patterning increases ecosystem resilience. However, these predictions rely on simplifying assumptions, such as assuming isotropic and infinitely large ecosystems, and empirical evidence directly linking spatial patterning to enhanced resilience remains scarce. We introduce a unifying framework, encompassing existing models for vegetation pattern formation in water-stressed ecosystems, that relaxes these assumptions. This framework incorporates finite vegetated areas surrounded by desert and anisotropic environmental conditions that lead to non-reciprocal plant interactions. Under these more realistic conditions, we identify a novel desertification mechanism, known as nonlinear convective instability in physics but largely overlooked in ecology. These instabilities form when non-reciprocal interactions destabilize the vegetation-desert interface and can trigger desertification fronts even under stress levels where isotropic models predict stability. Importantly, ecosystems exhibiting periodic vegetation patterns are more susceptible to nonlinear convective instabilities than those with homogeneous vegetation, suggesting that spatial patterning may reduce, rather than enhance, resilience. These findings challenge the prevailing view that self-organized patterning enhances ecosystem resilience and provide a new framework for investigating how spatial dynamics shape the stability and resilience of ecological systems under changing environmental conditions.

We consider a multitype Galton-Watson process that allows for the mutation and reversion of individual types in discrete and continuous time. In this setting, we explicitly compute the time evolution of quantities such as the mean and distributions of different types. This allows us in particular to estimate the proportions of different types in the long run, as well as the distribution of the first time of occurrence of a given type as the tree size or time increases. Our approach relies on the recursive computation of the joint distribution of types conditionally to the value of the total progeny. In comparison with the literature on related multitype models, we do not rely on approximations.
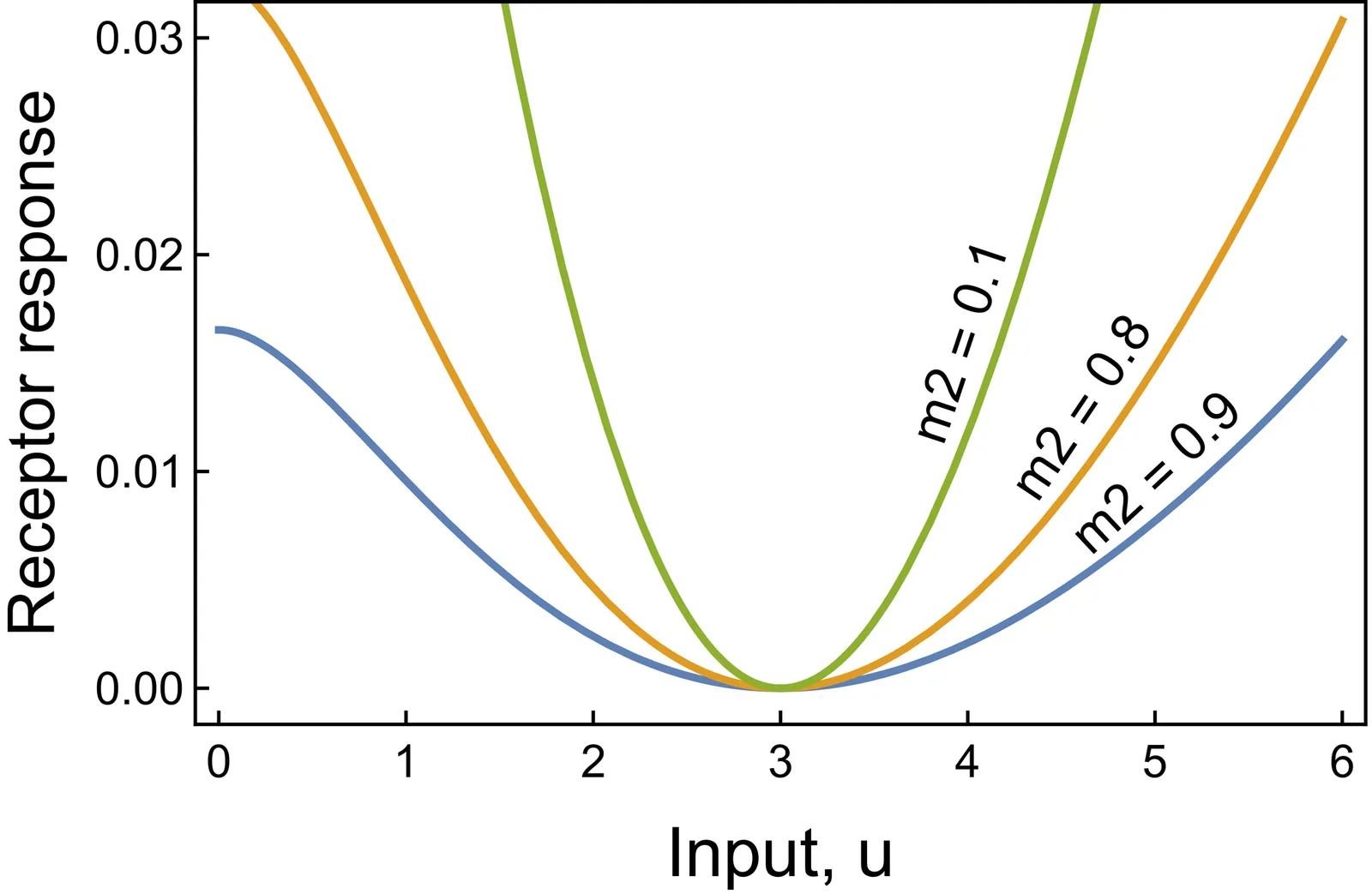
Anomaly detection is a well-established field in machine learning, identifying observations that deviate from typical patterns. The principles of anomaly detection could enhance our understanding of how biological systems recognize and respond to atypical environmental inputs. However, this approach has received limited attention in analyses of cellular and physiological circuits. This study builds on machine learning techniques -- such as dimensionality reduction, boosted decision trees, and anomaly classification -- to develop a conceptual framework for biological circuits. One problem is that machine learning circuits tend to be unrealistically large for use by cellular and physiological systems. I therefore focus on minimal circuits inspired by machine learning concepts, reduced to cellular scale. Through illustrative models, I demonstrate that small circuits can provide useful classification of anomalies. The analysis also shows how principles from machine learning -- such as temporal and atemporal anomaly detection, multivariate signal integration, and hierarchical decision-making cascades -- can inform hypotheses about the design and evolution of cellular circuits. This interdisciplinary approach enhances our understanding of cellular circuits and highlights the universal nature of computational strategies across biological and artificial systems.
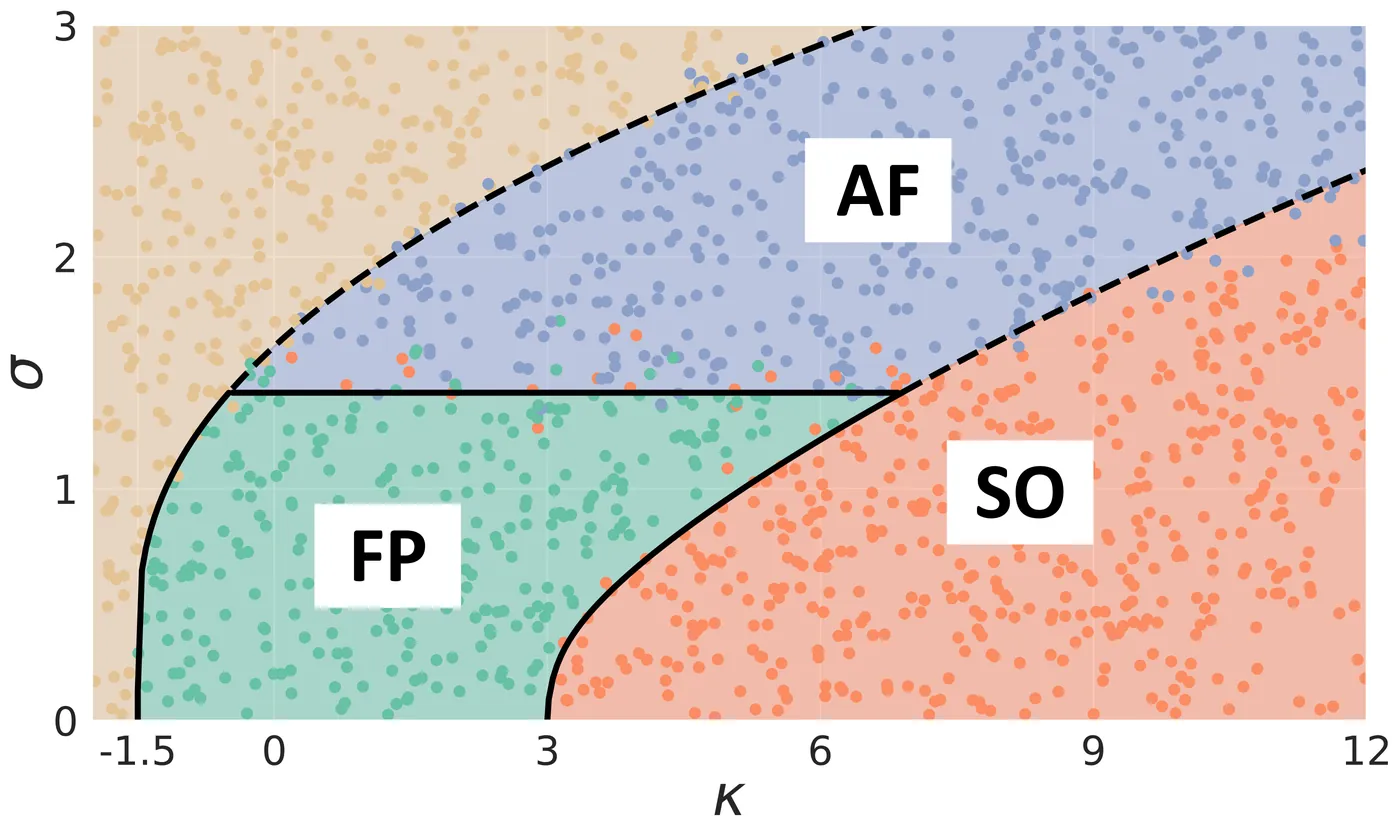
A central feature of complex systems is the relevance and entanglement of different levels of description. For instance, the dynamics of ecosystems can be alternatively described in terms of large ecological processes and classes of organisms, or of individual species and their relations. Low-dimensional heuristic 'macroscopic' models that are widely used to capture ecological relationships -- and commonly evidence out-of equilibrium regimes -- implicitly assume that species-level 'microscopic' heterogeneity can be neglected. Here, we address the stability of such macroscopic descriptions to the addition of disordered microscopic interactions. We find that increased heterogeneity can stabilize collective as well as species fluctuations -- contrary to the well-known destabilizing effect of disorder on fixed points. We analytically find the conditions for the existence of heterogeneity-driven equilibria, and relate their stability to a mismatch in microscopic time scales. This may shed light onto the empirical observation that many-species ecosystems often appear stable at aggregated levels despite highly diverse interactions and large fluctuations at the species level.
We consider a population model in which the season alternates between winter and summer, and individuals can acquire mutations either that are advantageous in the summer and disadvantageous in the winter, or vice versa. Also, we assume that individuals in the population can either be active or dormant, and that individuals can move between these two states. Dormant individuals do not reproduce but do not experience selective pressures. We show that, under certain conditions, over time we see two waves of adaptation. Some individuals repeatedly acquire mutations that are beneficial in the summer, while others repeatedly acquire mutations that are beneficial in the winter. Individuals can survive the season during which they are less fit by entering a dormant state. This result demonstrates that, for populations in fluctuating environments, dormancy has the potential to induce speciation.
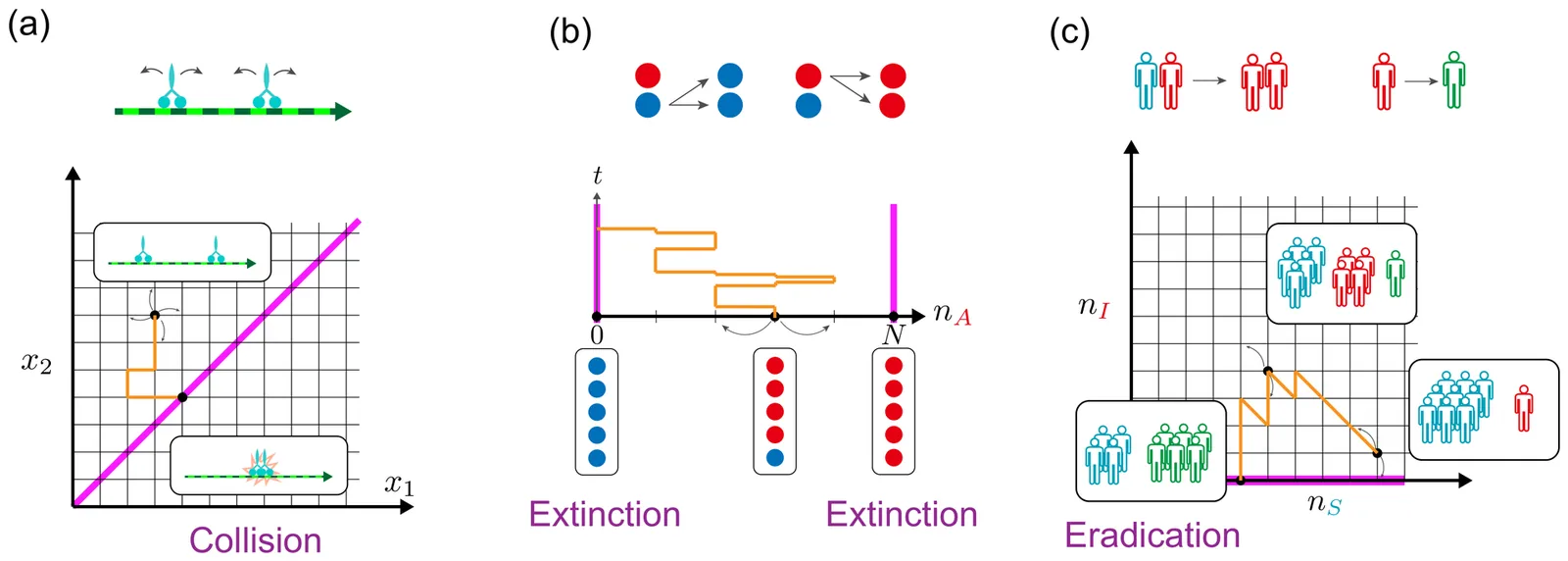
Controlling the stochastic dynamics of biological populations is a challenge that arises across various biological contexts. However, these dynamics are inherently nonlinear and involve a discrete state space, i.e., the number of molecules, cells, or organisms. Additionally, the possibility of extinction has a significant impact on both dynamics and control strategies, particularly when the population size is small. These factors hamper the direct application of conventional control theories to biological systems. To address these challenges, we formulate the optimal control problem for stochastic population dynamics by utilizing control cost functions based on the f-divergence, which naturally accounts for population-specific factors. If Kullback-Leibler (KL) divergence is adopted for the cost function, the complex nonlinear Hamilton-Jacobi-Bellman equation is simplified into a linear form, facilitating efficient computation of optimal solutions. We demonstrate the effectiveness of our approach by applying it to the control of interacting random walkers, Moran processes, and SIR models, and observe the mode-switching phenomena in the control strategies. Our approach provides new opportunities for applying control theory to a wide range of biological problems.
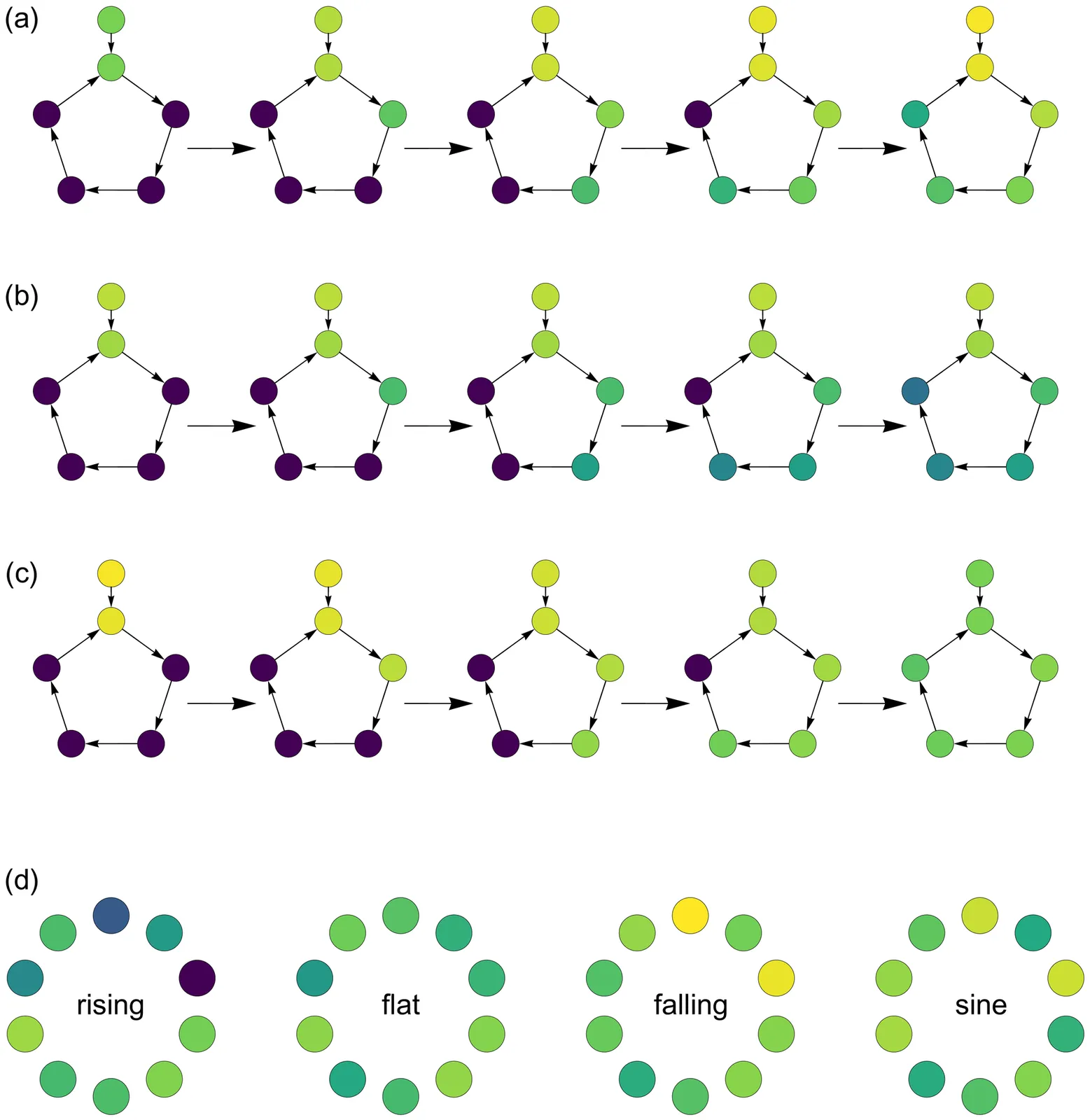
A biological circuit is a neural or biochemical cascade, taking inputs and producing outputs. How have biological circuits learned to solve environmental challenges over the history of life? The answer certainly follows Dobzhansky's famous quote that ``nothing in biology makes sense except in the light of evolution.'' But that quote leaves out the mechanistic basis by which natural selection's trial-and-error learning happens, which is exactly what we have to understand. How does the learning process that designs biological circuits actually work? How much insight can we gain about the form and function of biological circuits by studying the processes that have made those circuits? Because life's circuits must often solve the same problems as those faced by machine learning, such as environmental tracking, homeostatic control, dimensional reduction, or classification, we can begin by considering how machine learning designs computational circuits to solve problems. We can then ask: How much insight do those computational circuits provide about the design of biological circuits? How much does biology differ from computers in the particular circuit designs that it uses to solve problems? This article steps through two classic machine learning models to set the foundation for analyzing broad questions about the design of biological circuits. One insight is the surprising power of randomly connected networks. Another is the central role of internal models of the environment embedded within biological circuits, illustrated by a model of dimensional reduction and trend prediction. Overall, many challenges in biology have machine learning analogs, suggesting hypotheses about how biology's circuits are designed.

Muller's ratchet, in its prototype version, models a haploid, asexual population whose size~$N$ is constant over the generations. Slightly deleterious mutations are acquired along the lineages at a constant rate, and individuals carrying less mutations have a selective advantage. The classical variant considers {\it fitness proportional} selection, but other fitness schemes are conceivable as well. Inspired by the work of Etheridge et al. ([EPW09]) we propose a parameter scaling which fits well to the ``near-critical'' regime that was in the focus of [EPW09] (and in which the mutation-selection ratio diverges logarithmically as $N\to \infty$). Using a Moran model, we investigate the``rule of thumb'' given in [EPW09] for the click rate of the ``classical ratchet'' by putting it into the context of new results on the long-time evolution of the size of the best class of the ratchet with (binary) tournament selection, which (other than that of the classical ratchet) follows an autonomous dynamics up to the time of its extinction. In [GSW23] it was discovered that the tournament ratchet has a hierarchy of dual processes which can be constructed on top of an Ancestral Selection graph with a Poisson decoration. For a regime in which the mutation/selection-ratio remains bounded away from 1, this was used in [GSW23] to reveal the asymptotics of the click rates as well as that of the type frequency profile between clicks. We will describe how these ideas can be extended to the near-critical regime in which the mutation-selection ratio of the tournament ratchet converges to 1 as $N\to \infty$.
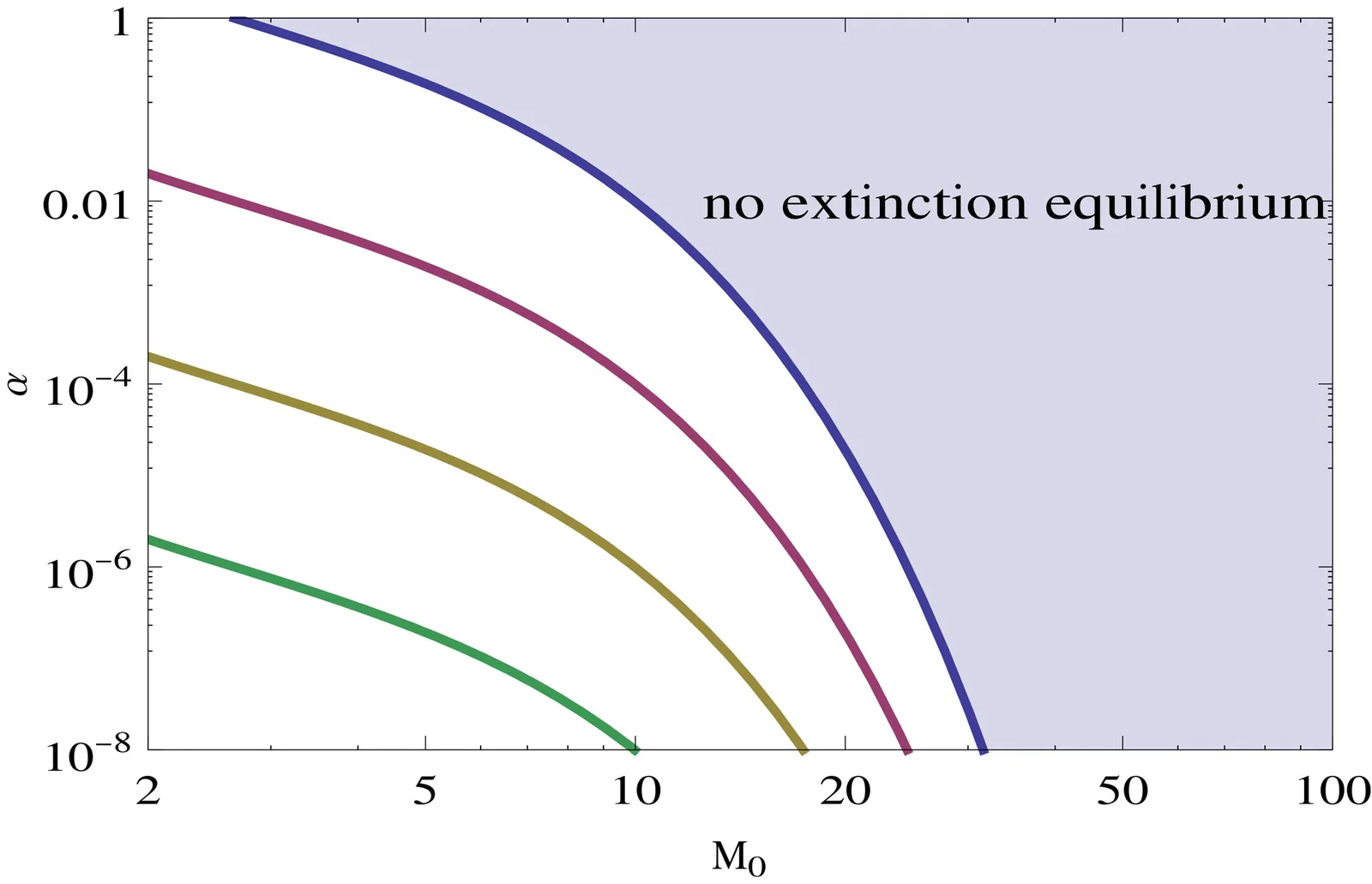
We investigate solutions to the TAP equation, a phenomenological implementation of the Theory of the Adjacent Possible. Several implementations of TAP are studied, with potential applications in a range of topics including economics, social sciences, environmental change, evolutionary biological systems, and the nature of physical laws. The generic behaviour is an extended plateau followed by a sharp explosive divergence. We find accurate analytic approximations for the blow-up time that we validate against numerical simulations, and explore the properties of the equation in the vicinity of equilibrium between innovation and extinction. A particular variant, the two-scale TAP model, replaces the initial plateau with a phase of exponential growth, a widening of the TAP equation phenomenology that may enable it to be applied in a wider range of contexts.
Reliable short term forecasting can provide potentially lifesaving insights into logistical planning, and in particular, into the optimal allocation of resources such as hospital staff and equipment. By reinterpreting COVID-19 daily cases in terms of candlesticks, we are able to apply some of the most popular stock market technical indicators to obtain predictive power over the course of the pandemics. By providing a quantitative assessment of MACD, RSI, and candlestick analyses, we show their statistical significance in making predictions for both stock market data and WHO COVID-19 data. In particular, we show the utility of this novel approach by considering the identification of the beginnings of subsequent waves of the pandemic. Finally, our new methods are used to assess whether current health policies are impacting the growth in new COVID-19 cases.
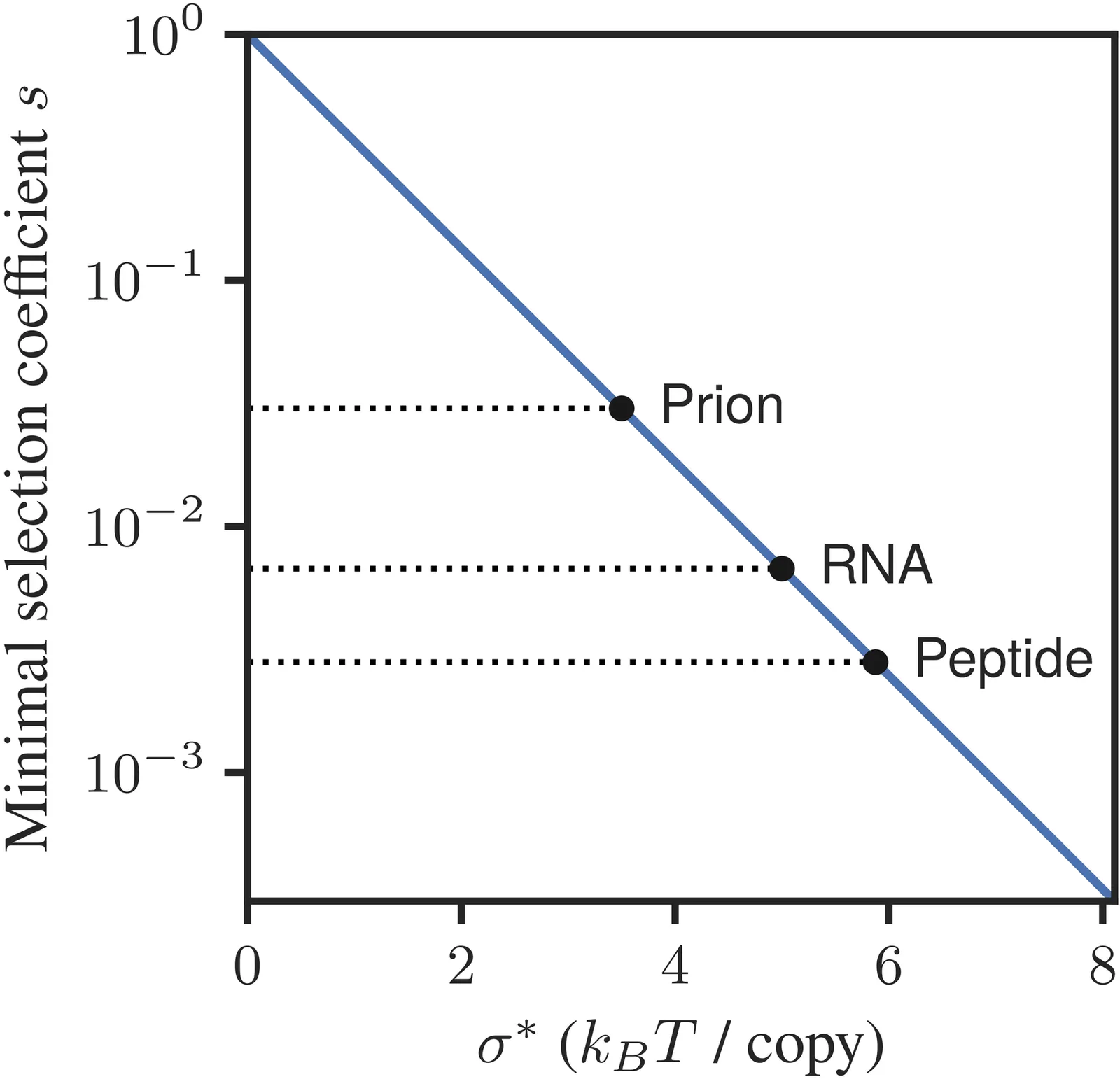
We consider the relationship between thermodynamics, fitness, and Darwinian selection in autocatalytic molecular replicators. We uncover a thermodynamic bound that relates fitness, replication rate, and thermodynamic affinity of replication. This bound applies to a broad range of systems, including elementary and non-elementary autocatalytic reactions, polymer-based replicators, and certain kinds of autocatalytic sets. In addition, we show that the critical selection coefficient (the minimal fitness difference visible to selection) is bounded by a simple function of the affinity. Our results imply fundamental thermodynamic bounds on selection strength in molecular evolution, complementary to other bounds that arise from finite population sizes and error thresholds. These bounds may be relevant for understanding thermodynamic constraints faced by early replicators at the origin of life. We illustrate our approach on several examples, including a classic model of replicators in a chemostat.